Echinoderm stereom gradient structures enable mechanoelectrical perception
TL;DR
Echinoderm stereom, a biomineralized cellular solid in sea urchin spines, exhibits mechanoelectrical perception with response potential and time exceeding echinoderm vision by orders of magnitude. This is due to gradient cellular structures along the spine axis, generating streaming potentials during liquid flow. Inspired by this, artificial structures show enhanced voltage output, enabling applications in underwater sensing.
Key Takeaways
- •Sea urchin spines have mechanoelectrical perception with response potential and time one to three orders of magnitude greater than echinoderm vision.
- •This perception arises from gradient cellular solids in stereom, creating differential charge density during liquid flow via streaming potentials.
- •Artificial spine-like structures, 3D-printed with gradient designs, show three-fold higher voltage output and eight-fold greater amplitude differential than gradient-free samples.
- •The findings advance understanding of load-sensitive biomimetic cellular solids, with potential for underwater spatiotemporal sensing and water resource utilization.
Tags
Abstract
Cellular solids ubiquitously exist in natural systems and are crucial for living organisms1,2. Their unique smooth branch and node morphologies are often seen as adaptations for enhanced mechanical performance3,4. Exploring alternative evolutionary functions can enrich the understanding of cellular solids, but it is frequently neglected. Here we show that the biomineralized cellular solids in echinoderm stereom (for example, sea urchin spine) have unexpected mechanoelectrical perception with response potential and response time, both of which are one to three orders of magnitude greater than those of echinoderm vision5. This exceptional perception originates from the gradient cellular solids (with varying void- or solid-phase diameters) along the [001] spine axis, generating a differential charge density across the stereom surface during liquid flow. Inspired by this natural wisdom, we create artificial spine-like structures using three-dimensional printing technology that exhibit three-fold higher voltage output and eight-fold greater amplitude differential than gradient-free samples, as well as a nature-inspired metamaterial mechanoreceptor capable of time-resolved self-monitoring information underwater. Our findings advance the understanding of load-sensitive biomimetic cellular solids (such as wood, sponge and trabecular bone), with the potential to develop functional gradient cellular materials towards underwater spatiotemporal sensing and water resource utilization.
Similar content being viewed by others

High strength and damage-tolerance in echinoderm stereom as a natural bicontinuous ceramic cellular solid
Structural and molecular distinctions of primary and secondary spines in the sea urchin Lytechinus variegatus
Toxigenic effects of sponges and benthic diatoms on marine invertebrates and their possible biotechnological applications
Main
Cellular solids (also known as structural foams1) are ubiquitous in natural and engineered systems, such as wood6, trabecular bone7, energy storage8 and catalysts9, owing to their excellent mass transport, mechanical properties and energy absorption. Recent efforts have been dedicated to developing cellular microarchitected materials inspired by natural systems (for example, echinoderm stereom) with extraordinary mechanical strength and damage tolerance3,10,11,12. These studies have shown the strengthening–toughening mechanism through branch damage bands and dual-scale hierarchical microlattices in cellular solids, inspiring the design of engineered architectures for both laboratory and industrial applications. It is notable that these natural cellular solids may not have primarily evolved for improved mechanical properties, but rather may be a by-product of the complex biomineralization process10. Uncovering previously unknown mechanisms beyond their traditionally recognized function of mechanical defence is essential for comprehensively understanding and using these naturally formed cellular solids.
Here we show a previously unexplored mechanoelectrical perception behaviour of the biomineralized spine of a long-spined sea urchin, Diadema setosum, which has a typical spike-like shape with a length of approximately 5–8 cm (Fig. 1a). In situ observations of a living D. setosum demonstrate that its spines have independent and highly responsive tactile perception abilities. On dropping a seawater droplet onto the apex of a spine, a swift and noticeable rotation of approximately 10° relative to the body shell axis occurs within 1 s. By contrast, the surrounding spines unexposed to the droplet exhibit no discernible response (Fig. 1b and Supplementary Video 1). The response time of the mechanoelectrical perception of the spine is approximately 88 ms, as measured using high-speed imaging (Supplementary Fig. 1 and Supplementary Video 2).
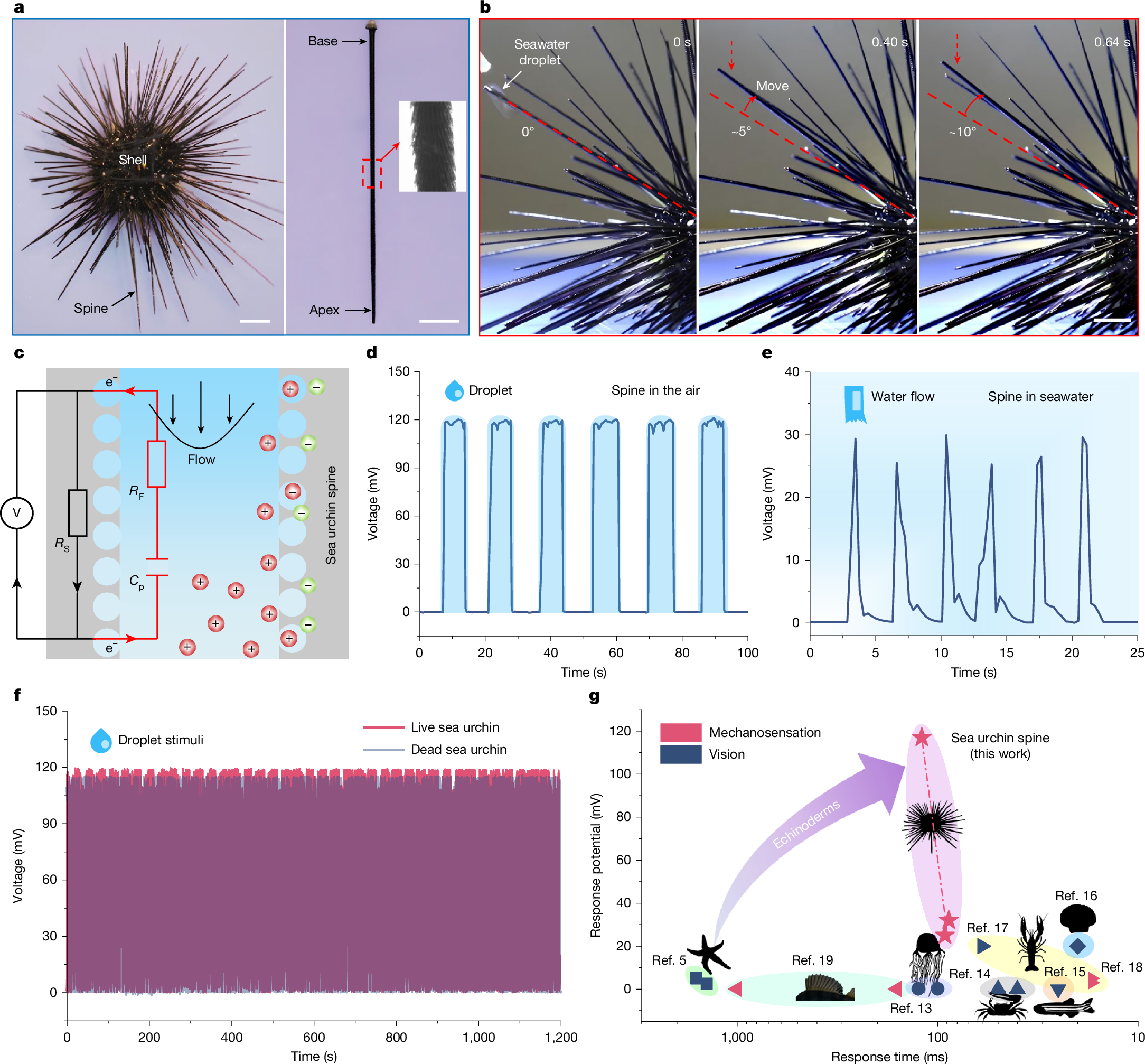
a, Optical images of a living D. setosum, showing a dome-shaped body shell surrounded by spike-like spines approximately 5–8 cm long. b, A droplet of seawater falls onto a spine apex (red arrows), resulting in a rotation of about 10° within 1 s. c, Schematic of electrical measurement on the sea urchin spine. When water flows through microchannels, the spines are negatively charged, resulting in a positively charged surrounding fluid. d, Response voltage across the spine in air during droplet stimulus, reaching a maximum amplitude of around 116 mV. e, Voltage measurements from the submerged spine, showing a transient response to seawater flow stimulations. f, Voltage output of the same sea urchin spine under droplet stimulus, showing no substantial difference between live and dead states. g, Comparison of the response potential and response time of some typical marine animals5,13,14,15,16,17,18,19. Scale bars, 2 cm (a, left), 1 cm (a, right,b).
Mechanoelectrical perception ability
We use a data acquisition and digital multimeter system to monitor the haptic-sensing electrical event in the spine of a living D. setosum, under precisely controlled hydration stimulations delivered by a custom seawater droplet and flow generator (Supplementary Fig. 2). A schematic of the electrical measurement on the live sea urchin spine is shown in Fig. 1c. Conductive electrodes are connected to two different locations along the spine to measure the voltage difference. Sea urchin spines exposed to air show no detectable initial electrical potential without external stimulation. On the seawater droplet dropping onto the spine apex, it exhibits an electrical potential with a peak amplitude of about 116 mV (Fig. 1d and Supplementary Video 3). This potential rapidly declines towards baseline after droplet removal but recovers instantly to peak levels on subsequent droplet arrival, indicating real-time responsiveness to the droplet stimulus. A similar electrical response with a peak potential of about 30 mV is detectable in the submerged spine during seawater flow stimulation (Fig. 1e and Supplementary Video 4). Notably, the response potential to droplet stimulus shows no substantial difference between living and dead sea urchin spines (Fig. 1f), consistent with histological evidence confirming the absence of viable cellular tissue on the spine exterior surface or within the three-dimensional (3D) structure of the stereom (Supplementary Fig. 3 and Supplementary Note 1). These results indicate that the response potential of the spine to droplet stimulation seems independent of living tissue, revealing a previously unrecognized mechanism, discussed in more detail later. This unexpected mechanoelectrical perception demonstrates potential amplitudes and response time one to three orders of magnitude greater than those of echinoderm vision5. It also surpasses the perceptual abilities observed in some common marine animals, such as box jellyfish, fiddler crabs and zebrafish13,14,15,16,17,18,19 (Fig. 1g and Supplementary Table 1). To the best of our knowledge, echinoderms lack specialized sensory organs20. Thus, the mechanoelectrical perception of sea urchin spines and their underlying mechanisms remain unexplained.
Gradient cellular structures in spines
Scanning electron microscopy (SEM) and micro-computed tomography (μ-CT) of the fractured spine show their continuous morphological characteristics along the [001] direction, spanning from the spine base to the apex (Fig. 2a–d). The D. setosum spine is primarily composed of three interconnected parts: a porous stereom network, an interior hollow channel and a dense outer ratchet layer (Fig. 2e). The stereom network comprises open-cell microstructures composed of magnesium-containing calcite, amorphous calcium carbonate (Supplementary Figs. 4 and 5) and a minor fraction of intracrystalline organics (approximately 1.4 wt%)21. It presents a typical bicontinuous morphology (including solid and void phases22) with a highly curved and smooth minimum surface10,23. Notably, both the solid and void phases exhibit a notable diameter gradient along the [001] direction within the spine.
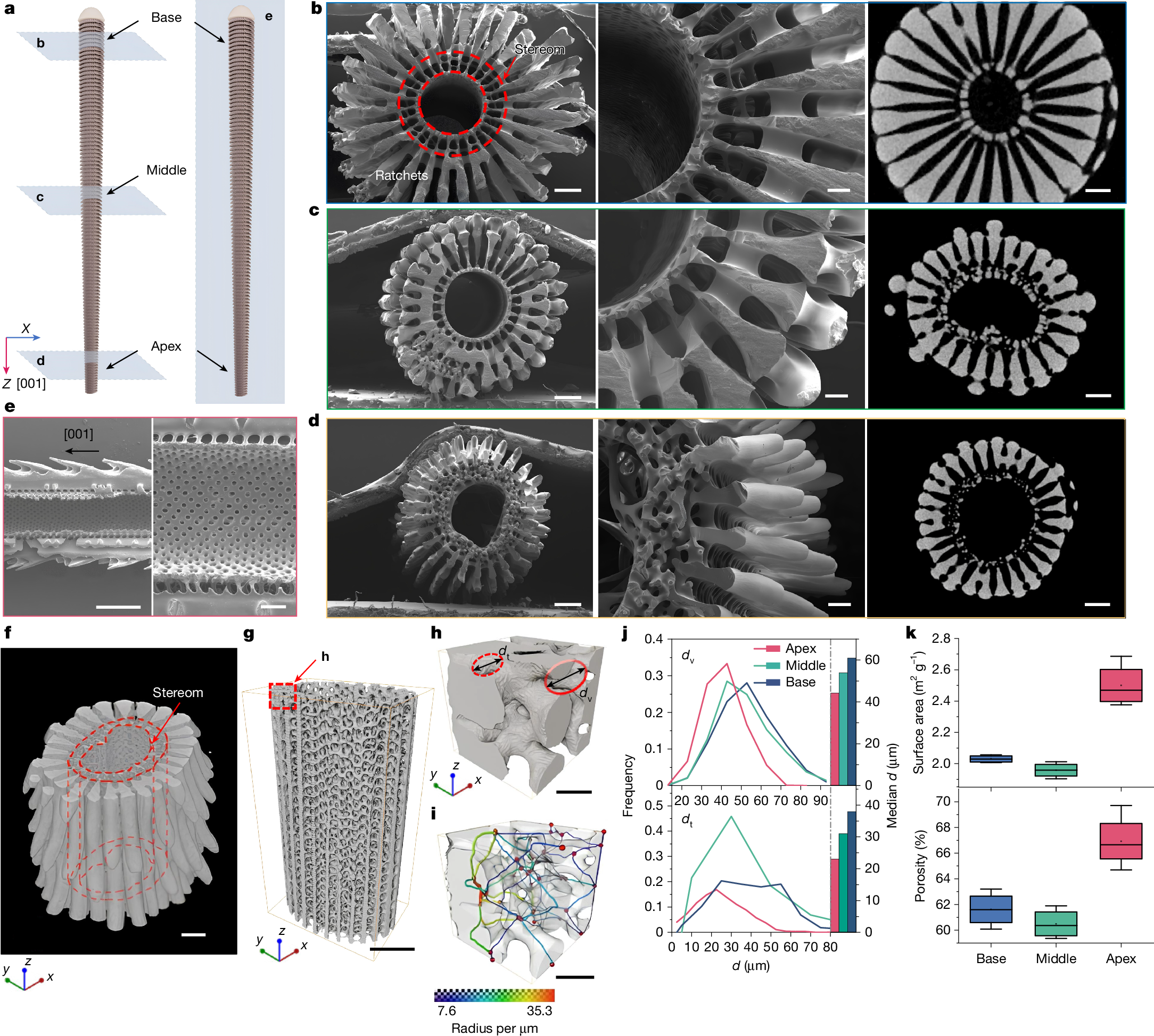
a, Schematic of 3D spine models with cross-sections along the transverse (left) and longitudinal (right) directions. b–d, SEM and μ-CT images of the transverse cross-section at the spine base, middle and apex. The stereom network exhibits a notable diameter gradient along the [001] direction. e, SEM images of the longitudinal cross-section of the spine, showing a highly interconnected cellular network. f–g, µ-CT reconstructions and the corresponding stereom network at the middle portion. h–i, Representative 3D cellular structure of stereom network and the corresponding void phase with throats coloured by their diameters. dv and dt represent the diameters of the void phase and throats, respectively. j, Distribution and median values of dv and dt, showing a slow decline along the [001] direction. k, Specific surface area and porosity of stereom microstructure within different spine portions. Scale bars, 200 μm (b–d, left and right); 100 μm (b–d, middle); 500 μm (e, left); 200 μm (e, right); 200 μm (f,g); 50 μm (h,i). In k, the small open square represents mean; centre line, median; box limits, upper and lower quartiles; whiskers, minimum and maximum; n = 4 samples.
µ-CT reconstruction of the D. setosum spine shows the intricate details of the solid- and void-phase morphologies within the stereom networks (Fig. 2f–i, Supplementary Fig. 6 and Supplementary Video 5). According to the volume-based local thickness definition24, the diameter of the largest sphere that can be fully embedded in the void phase is defined as the local thickness dv, whereas the thinnest region (constriction) in the void space is defined as the throat25 with diameter dt (Fig. 2h,i). At the spine base, dv ranges from 25 μm to 95 μm with a median of about 60.9 μm. Meanwhile, at the spine apex, dv ranges from 15 μm to 75 μm with a median of about 44.2 μm. As a result, both dv and dt of the stereom microstructure decrease gradually along the [001] direction (Fig. 2j and Supplementary Figs. 7–9). This notable stereom gradient along the [001] direction could facilitate internal fluid convection and transport within the spine. Moreover, the stereom microstructure at the spine apex exhibits a remarkable specific surface area of 2.50 ± 0.14 m2 g−1 and a porosity of 66.9 ± 2.1%, which are approximately 23% and 5.3% higher than those at the spine base, respectively (Fig. 2k). The increased porosity and minimal void-phase size of the stereom microstructure at the spine apex could enhance the interfacial interactions between solids and liquids, whereas the expanded specific surface area could offer an abundance of sites for interfacial collisions.
Mechanoelectrical perception mechanism
Contact angle measurements show a wettability difference on the spine surface along the [001] direction (Supplementary Note 2 and Supplementary Fig. 10), suggesting that liquid can readily penetrate the stereom network at the spine bases and then progress toward the apex. To simulate this process and detect mechanoelectrical perception in vitro, we perform the electrical test by injecting liquids into the stereom network from its base using a microlitre syringe. On complete water diffusion across the spine, an immediate potential exceeding 20 mV is detected (Fig. 3a, Supplementary Figs. 11 and 12 and Supplementary Video 6). Quasi-static piezoelectric constant measurements show no detectable piezoelectricity within spines (Supplementary Fig. 13). In the fully wetted state, the spine exhibits a real-time potential response to water flow, generating a measurable voltage during fluid motion that dissipates on flow cessation (Fig. 3b). These findings indicate that the measured potential during flow originates from streaming potential26,27. The initial spine–liquid contact induces interfacial charge transfer and establishes an electric double layer (EDL)28. When the spine is fully wetted, liquid flow shears the EDL, inducing charge separation and rearrangement that generate a streaming potential. On flow cessation, charge separation terminates, enabling charge redistribution and neutralization that dissipates the potential difference (Fig. 3c, Supplementary Fig. 14 and Supplementary Note 3). This streaming potential is also detected during seawater flow through the spine. Zeta potential (ζ) tests show that increased ionic strength in seawater induces the EDL compaction, reducing interfacial ion mobility and charge density as per Gouy–Chapman theory29 (Fig. 3d).
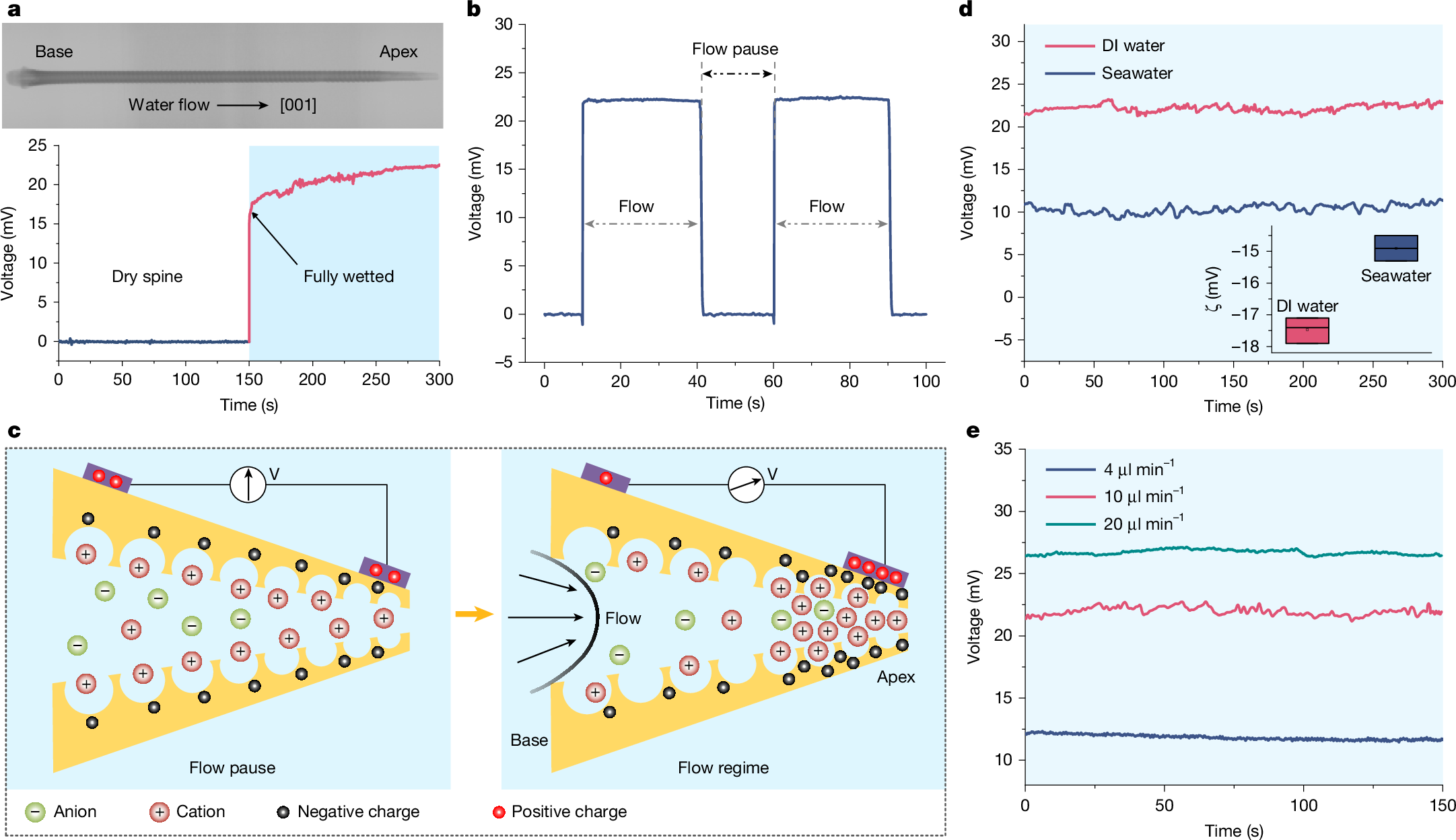
a, Voltage output of the spine during water flows, showing an immediate potential exceeding 20 mV on complete water diffusion across the spine. b, Voltage measurement from the fully wetted spine, showing a real-time response to water flow. c, Schematic of streaming potential generation during liquid flow, showing a substantial interfacial charge separation. At the spine apex, the increased specific surface area in stereom promotes EDL formation and collision frequency, enhancing interfacial charge density. d, Streaming potential of the spine during deionized (DI) water and seawater injection (inset, ζ of the spine in DI water compared with seawater). High ionic concentration in seawater induces the EDL compaction, reducing interfacial ion mobility and charge density. e, Voltage output of the spine at different flow velocities, showing an increased streaming potential with flow velocity. In the inset of d, dot represents mean; centre line, median; box limits, upper and lower quartiles; n = 3 samples.
Within sea urchin spines, the gradient cellular structure induces differences in fluid velocity and liquid pressure across the stereom surface under liquid flows, as demonstrated by finite element simulations of a simplified 3D symmetric model (Supplementary Fig. 15 and Supplementary Note 3). Compared with the spine base, the stereom microstructure at the spine apex features reduced void-phase diameters that en