A blastoporal organizer in a ctenophore
Abstract
In an iconic experiment in 1924, Hilde Mangold and Hans Spemann established that the dorsal blastopore lip of amphibian embryos functions as an organizer and induces a secondary body axis when transplanted into a host embryo1. This discovery demonstrated that specific embryonic regions can regulate embryonic patterning and lead to the establishment of an entire body axis. Subsequent studies have revealed that cnidarians, the sister group to Bilateria, also possess a blastoporal embryonic organizer2,3. However, the evolutionary origin of the organizer remains unclear. Here we report that the blastopore lip of the ctenophore Mnemiopsis leidyi, a member of the evolutionary sister group to all other metazoans4,5, exhibits organizer activity. We show that transplanted fragments of blastopore lip tissue from M. leidyi gastrula induce secondary pharynx and mouth formation. Moreover, transphyletic transplantation experiments show that the blastopore lip of M. leidyi leads to the generation of a secondary body axis in embryos of the cnidarian Nematostella vectensis. Organizer function in M. leidyi requires both β-catenin and TGFβ signalling, and the TGFβ-family ligands probably provide this inductive capacity. These findings reveal the deep homology of the blastoporal organizer in ctenophores, cnidarians and vertebrates, implying the ancestral organizer role of the blastopore lip. We propose that the emergence of the organizer was an essential innovation that facilitated the change from the temporal cell differentiation of unicellular relatives to the spatial cell differentiation of the first multicellular embryo.
Main
The concepts of the embryonic organizer and induction were introduced and developed by Hilde Mangold and Hans Spemann based on their landmark experiments in the 1920s1,6. Their studies on transplantation of the dorsal blastopore lip of amphibian embryos demonstrated that this tissue has the capacity to induce the formation of a secondary body axis by recruiting neighbouring host cells (Fig. 1a). The secondary axis is formed through interactions between the transplanted cells and the host embryo tissues, and the data highlighted the critical role of the organizer in directing host tissue differentiation and morphogenesis. This experiment provided clear evidence that specific cell populations in embryos can influence the differentiation of neighbouring cells. Subsequent comparative evolutionary studies revealed the presence of an embryonic organizer in a non-bilaterian animal, the cnidarian N. vectensis2,3. Transplanting a section of the blastopore lip from an early gastrula of N. vectensis to another embryo at the same stage of development results in the induction of a secondary oral–aboral axis (Fig. 1b). The central role of WNT–β-catenin signalling in driving the organizing activity of cnidarian and vertebrate blastopore lips suggest the potential homology of these structures. Despite extensive research on the embryonic organizer in bilaterians, particularly chordates (the zebrafish Danio rerio and the frog Xenopus laevis)7,8, and their sister group cnidarians (N. vectensis)2,3, the evolutionary history of the embryonic organizer remains unsolved. It is still unclear whether embryonic organizers exist in other non-bilaterian animals predating the cnidaria–bilaterian separation, like sponges (Porifera) and comb jellies (Ctenophora) (Fig. 1c). Ctenophores are of particular interest because they are considered to occupy a key phylogenetic position as a sister group to all other multicellular animals4,5. A ctenophore embryonic organizer, if homologous to those in cnidarians and bilaterians, would suggest that an organizer emerged alongside the advent of multicellularity.
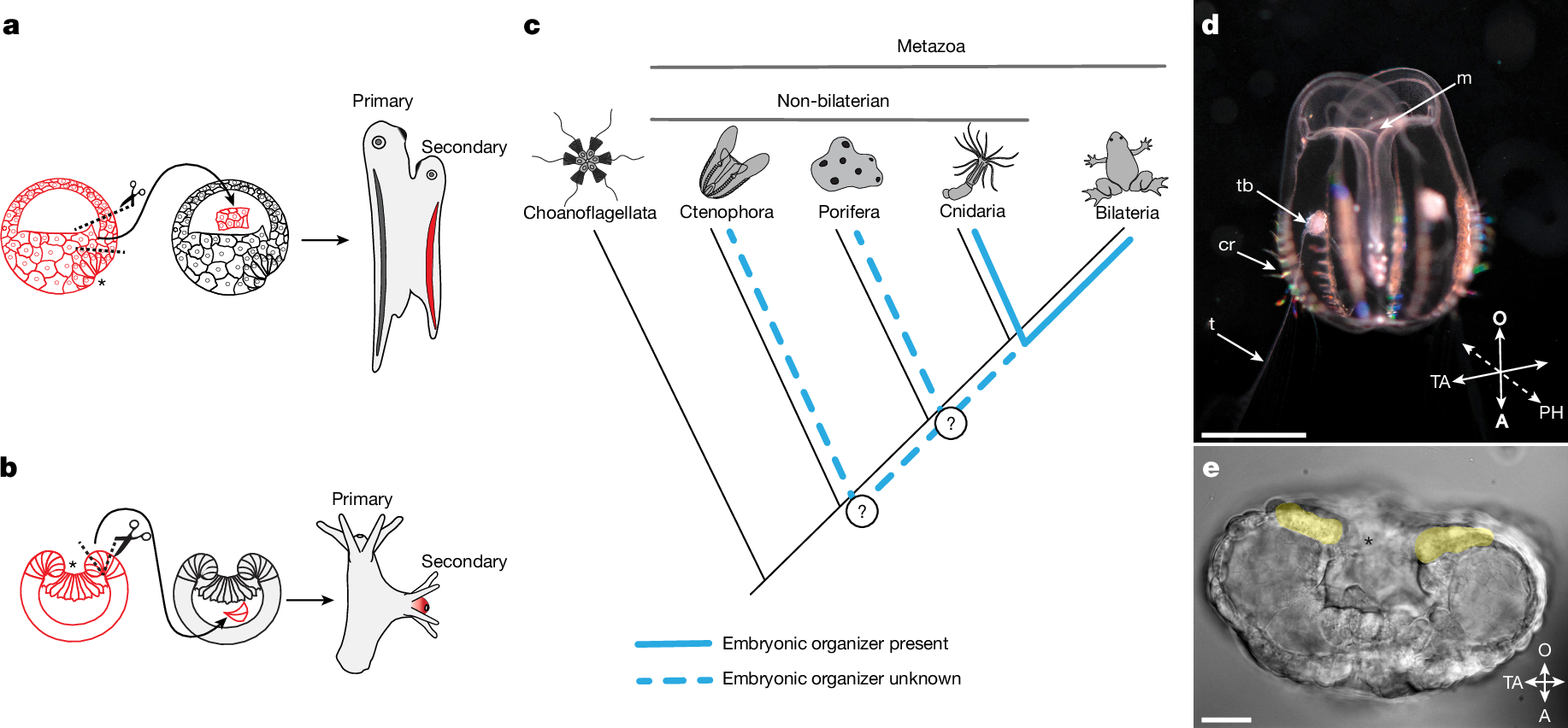
a,b, Schematics of embryonic organizer transplantation in amphibians (a) and cnidarians (anthozoans) (b). In both cases, transplantation of the blastopore lip fragment from one embryo to another induces a secondary axis involving host tissue. c, Presence of an embryonic organizer in different metazoan groups. The embryonic organizer has so far been described and studied only in bilaterians and their sister group cnidarians. However, it remains unknown whether an embryonic organizer exists in Porifera or Ctenophora; therefore, it is unclear when the embryonic organizer emerged in animal evolution. d, Cydippid stage of M. leidyi (Ctenophora). Scale bar, 2.5 mm. e, M. leidyi embryo at the gastrula stage. Cells surrounding the blastopore are highlighted yellow. Scale bar, 20 μm. Images in d and e are representative images from normal wild-type M. leidyi at the indicated developmental stages. Such normal morphology was consistently observed in all examined specimens. Asterisks (a,b,e) indicate the blastopore. A, aboral; cr, comb row; m, mouth; O, oral; PH, pharyngeal axis; t, tentacle; TA, tentacular axis; tb, tentacle bud.
Ctenophore blastopore lip is an organizer
To analyse the inductive capacity of different parts of the M. leidyi gastrula embryo, we transplanted blastopore lip tissue and lateral ectoderm to host embryos of the same stage of development (Fig. 2a and Extended Data Fig. 1). In our experiments, 41.8% (28 out of 67) of blastopore lip transplantations resulted in the formation of a complete secondary pharynx with a mouth opening (Fig. 2b). Moreover, 14.9% (10 out of 67) of embryos formed a mouth-like structure without a developed pharynx (Fig. 2c), and 43.3% (29 out of 67) developed normally (Fig. 2d). We did not observe duplication of any other anatomical structures. By contrast, embryos that received lateral ectoderm grafts (n = 49) never formed secondary pharynxes or mouth openings. Similarly, after control incisions without grafting (n = 63), we did not observe the formation of any additional structures (Fig. 2e–h). In cases when the ectopic pharynx was fully developed, the primary and secondary induced pharynxes fused together before connecting to the ‘stomach’ (infundibulum) and merging into a single digestive system (Fig. 2i,j). Both the primary and the ectopically induced mouths were fully functional and capable of capturing food, such as rotifers, and delivering it into the shared gastrovascular system (Fig. 2k).
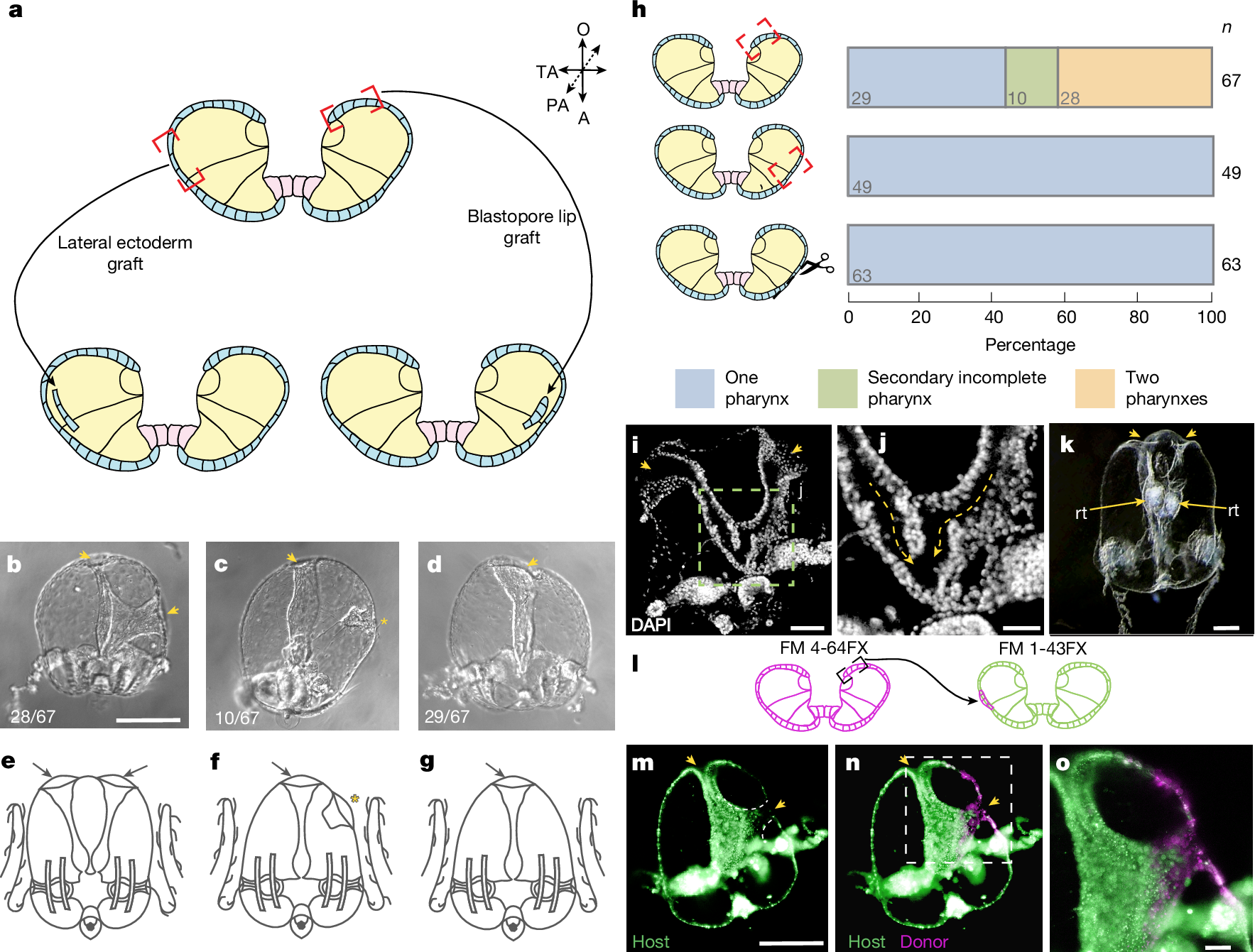
a, Schematic of the grafting experiment: transplantation of the blastoporal lip tissue or transplantation of lateral ectoderm. b–g, Images (top row) and schematics (bottom row) of the phenotypes obtained in transplantation experiment: two pharynxes (b,e), a secondary incomplete pharynx (c,f) or one pharynx (d,g). Arrowheads indicate a mouth opening with a fully formed pharynx in the cydippid. The asterisk indicates the formation of a mouth-like structure without a pharynx. Numbers in the bottom left present the fraction of embryos displaying this phenotype in the blastoporal lip tissue transplantation experiment. h, Transplantation of only the blastopore lip of a M. leidyi embryo can induce ectopic development of the fully formed axial structures or an ectopic secondary incomplete axis (top). By contrast, transplantation of the lateral ectoderm (middle) or incision without transplantation (bottom) results in the formation of a cydippid with only one body axis. Stacked bars show pooled proportions. i,j, Images of two fully formed pharynxes fused distally towards the infundibulum (n = 3). i, Arrowheads indicate the mouth openings. j, Arrows indicate the direction of fusion of the two pharynxes. k, Two fully developed pharynxes are visible, each with an ingested rotifer (rt). Arrowheads denote the mouth openings, and arrows indicate rotifers located in the pharyngeal cavities (n = 4). l, Schematic of the experiment for transplanting M. leidyi blastopore lip labelled with the vital dye FM4-64FX into a host embryo labelled with FM1-43FX. m–o, Images showing host tissues (m), and how the grafted blastopore contributed to the formation of an ectopic pharynx with integration of host tissues (n); close-up of the ectopic pharynx and mouth (o) (n = 5). Arrowheads denote the mouth openings. Scale bars, 250 μm (b–d,m,n), 100 μm (k), 50 μm (i,o), 25 μm (j).
Organizer transplants recruit host tissue
An essential characteristic of an organizer is to recruit surrounding host tissues to form induced structures after transplantation1,9. To explore this effect in our experimental model, we transplanted pieces of the blastopore lip, which were labelled with the vital membrane dye FM4-64FX, into FM1-43FX-labelled host gastrula embryos (Fig. 2l and Extended Data Fig. 2a). In transplantations that resulted in the induction of a fully developed ectopic pharynx, the pharyngeal tissue was composed of a mix of both host cells (FM1-43FX-labelled) and transplanted cells (FM4-64FX-labelled) (Fig. 2m–o).
However, the distribution of donor and host cells in the induced pharynx varied. In one instance, the ectopic pharynx consisted of donor and host tissues with a sharp boundary between them (Extended Data Fig. 2b–d). Alternatively, only one wall of the ectopic pharynx contained host-derived cells (Extended Data Fig. 2e–g) or the induced pharynx was composed of a uniformly mixed population of donor and host cells (Extended Data Fig. 2h–j). Notably, host cells were never observed in the area of the ectopic mouth opening (Fig. 2m–o and Extended Data Fig. 2b–j). This kind of split between donor and host contributions is also seen in classical Mangold–Spemann grafts. That is, donor cells largely contribute to the axial midline (for example, the ectopic notochord is almost entirely donor-derived), whereas surrounding parts of the induced axis often contain a mixture of donor and host cells1. Here we determined that the phenotype described as a ‘mouth-like structure without a developed pharynx’ (Fig. 2c,f) results from failed induction, as this structure is composed exclusively of donor tissue (Extended Data Fig. 2k–m).
For transplantations in which the blastopore lip graft did not induce an ectopic pharynx, we selectively labelled donor cells with FM4-64FX to trace the distribution of the transplanted tissues (Extended Data Fig. 3). In these embryos, we observed considerable variability in the distribution of donor-derived tissue. Donor cells were incorporated entirely in the primary pharynx (Extended Data Fig. 3d–f), scattered across the superficial epithelium (Extended Data Fig. 3g–i) or integrated into the tentacle bulb and tentacles (Extended Data Fig. 3g–l).
Thus, the formation of the secondary pharynx and mouth following blastopore lip transplantation in ctenophores involves the recruitment of host cells into the ectopically developing structure, which is a hallmark of a bona fide organizer.
Transphyletic axis induction
As our experiments confirmed that the ctenophore blastopore lip is an embryonic axial organizer, we investigated whether the M. leidyi blastopore lip can induce axis formation in other animals. We transplanted the blastopore lip from M. leidyi gastrula into the blastocoel of embryos of the sea anemone N. vectensis (Cnidaria) (Fig. 3a). We assessed the inductive capacity of the blastopore lip by analysing the expression of N. vectensis FoxA (NvFoxA; throughout, the first two letters prefixing a gene or protein indicate the species), a pharyngeal marker, which is consistently associated with the formation of a secondary axis during induction in N. vectensis2. As M. leidyi lacks a FoxA orthologue10, the specific NvFoxA probe used in the xenotransplantation assay can only detect NvFoxA-expressing N. vectensis cells. Thus, an NvFoxA-positive signal in the ectopic axis reflects the recruitment and pharyngeal specification of N. vectensis cells in the secondary axis induced by the ctenophore organizer. We observed ectopic NvFoxA expression in 15.7% (11 out of 70) of the transplantations (Fig. 3b–f). No ectopic NvFoxA expression was detected in any N. vectensis embryos grafted with M. leidyi lateral ectoderm (n = 53 total transplantations; Fig. 3f). Therefore, the embryonic organizer of ctenophores is capable of inducing an ectopic axis in a cnidarian, despite the deep evolutionary divergence of these two animal lineages. Therefore, similar mechanisms responsible for embryonic induction in Ctenophora must also be present in Cnidaria.
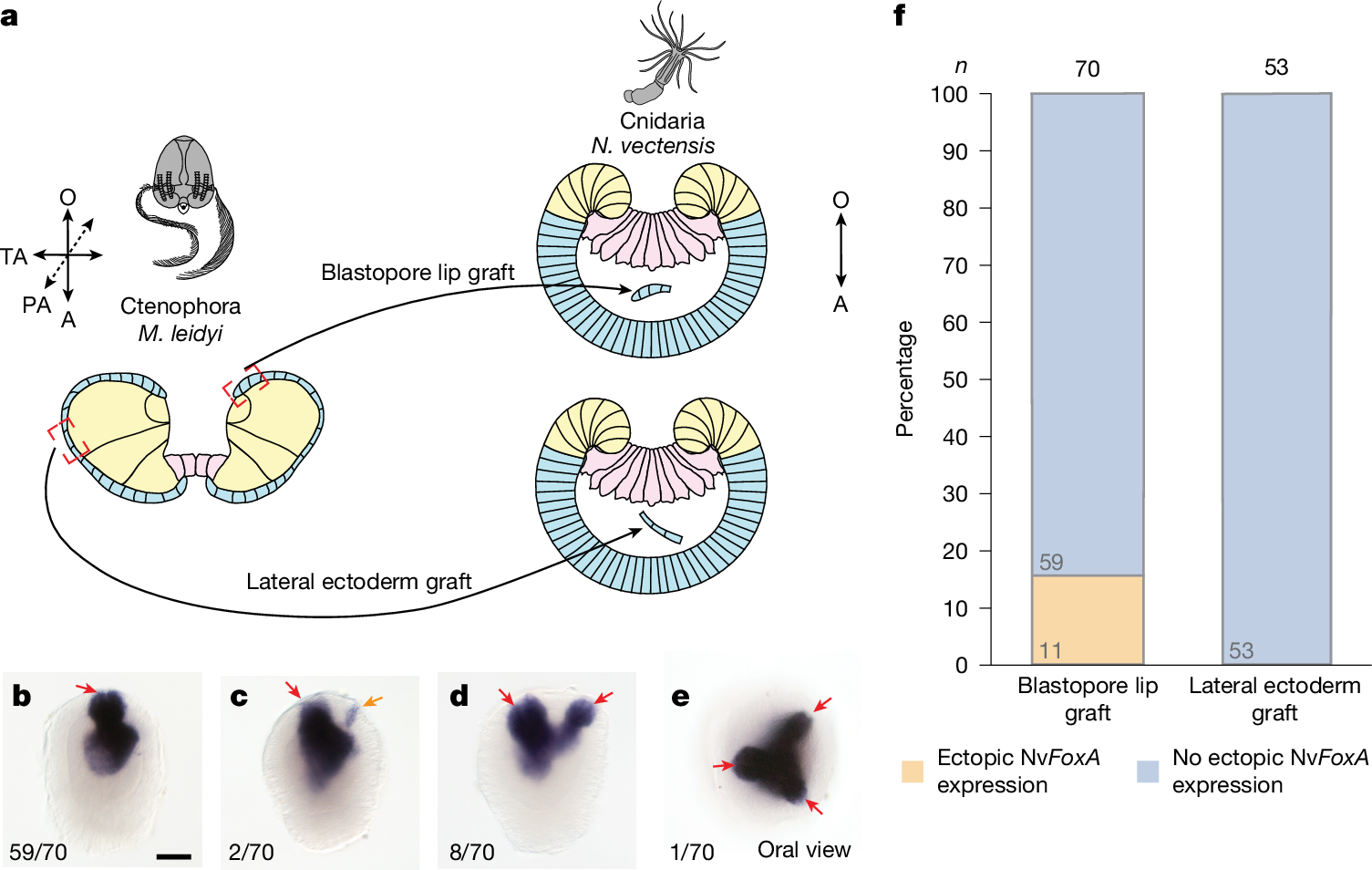
a, Schematic of the xenotransplantation experiment for grafting M. leidyi gastrula tissues into N. vectensis gastrula embryos. b–e, Images of phenotypes obtained in the xenotransplantation experiment, which analysed N. vectensis planula NvFoxA expression after M. leidyi organizer xenotransplantation: no ectopic expression (b) or ectopic expression associated with no distinct pharynx (c), one morphologically distinct pharynx (d) or two morphologically distinct pharynxes (e). The expression of the pharyngeal marker gene NvFoxA in N. vectensis embryos after transplantation of M. leidyi blastopore lip indicates the formation of multiple body axes. Red arrows indicate NvFoxA expression associated with morphologically distinct mouth opening formation (b–e), whereas the orange arrow indicates ectopic expression without such association (c). Numbers in the bottom left present the fraction of embryos displaying this phenotype. Scale bar, 50 μm. f, Quantification of transplantation of the blastopore lip of M. leidyi gastrula, but not lateral ectoderm, can induce the ectopic expression of NvFoxA in N. vectensis gastrula. Stacked bars show pooled proportions.
A reciprocal N. vectensis to M. leidyi blastopore lip transplantation would strengthen our argument that the molecular basis of axis induction is shared across Metazoa. However, such an experiment is not feasible for two reasons. First, embryonic cells of M. leidyi and N. vectensis show essentially no adhesion to each other. Second, M. leidyi embryos do not form a blastocoel at any stage of embryonic development that would enable stable retention of a heterologous donor graft. By contrast, the presence of a blastocoel in N. vectensis embryos allows stable retention of donor tissue transplanted from M. leidyi.
β-Catenin and TGFβ specify the organizer
To characterize the inductive nature of the ctenophore organizer, we investigated potential signalling pathways that are involved in the organizer region in M. leidyi. β-Catenin signalling influences primary axis polarity both in bilaterian animals11 and in non-bilaterian animals (for example, N. vectensis, Clytia hemisphaerica and Dynamena pumila)3,12,13. Therefore, its role in axial patterning probably predates the emergence of bilaterally symmetric animals. We hypothesized that the β-catenin pathway represents a universal signalling pathway that underlies the function of the embryonic organizer in all multicellular animals, including ctenophores.
To determine the contribution of β-catenin signalling to the organizer and oral structure development in M. leidyi, we treated embryos with the β-catenin pathway inhibitor iCRT14 and the activator CHIR99021. Embryos treated with 10 μM iCRT14 from the 2–4-cell stage to the end of gastrulation (8 h after the first division) (Extended Data Fig. 4a,b) developed into cydippid larvae, of which 67.5% (85 out of 126) exhibited a shortened pharynx and tentacle defects (Extended Data Fig. 4e,f). This phenotype closely recapitulated that observed in embryos of M. leidyi with Brachyury (MlBrachyury) knocked down or knocked out14,15. Inhibition of β-catenin signalling with iCRT14 also strongly reduced MlBrachyury expression (Extended Data Fig. 4g). In normal development, Brachyury is expressed during gastrulation in ectodermal cells surrounding the blastopore and continues to be expressed in the developing pharynx16. In M. leidyi, expression of the ancestral organizer-associated gene MlLhx1/5, as well as the aboral marker MlLhx3/4 (ref. 17), remained unchanged (Extended Data Fig. 4h,i).
Treatment with the β-catenin activator CHIR99021 (2.5 μM) from the 2–4-cell stage to 8 h after the first division (Extended Data Fig. 4j,k) did not lead to any visible morphological changes (Extended Data Fig. 4l,m), nor did it alter the expression of MlBrachyury, MlLhx1/5 or MlLhx3/4 (Extended Data Fig. 4n). Extending the treatment to 24 h after the first division, however, resulted in clear phenotypic effects by 2 days after fertilization (Extended Data Fig. 4o–r). Specifically, 19.4% of larvae (26 out of 134) developed a markedly expanded pharynx (Extended Data Fig. 4q), and 5.2% (7 out of 134) formed two closely adjacent mouth openings (Extended Data Fig. 4r). In the latter cases, the two mouths were positioned close to each other, which was probably due to the separation of the abnormally expanded pharynx by prolonged β-catenin activation. Increasing the concentration of CHIR99021 or prolonging exposure did not enhance these phenotypes but instead caused developmental arrest and embryo death. Previous work similarly reported minimal effects on early M. leidyi development after β-catenin signalling activation using compounds such as LiCl or alsterpaullone18. The fact that blocking β-catenin signalling in M. leidyi inhibits MlBrachyury expression, whereas activation of the pathway at early developmental stages has no detectable effect, suggests that β-catenin is necessary for MlBrachyury expression but not sufficient to activate expression or expand the MlBrachyury-positive domain.
It has previously been demonstrated that blocking the TGFβ–SMAD2/3 signalling cascade with SB431542, an inhibitor of the TGFβ type I receptor (ALK4/5/7), disrupts development of the ctenophore organizer region, leading to pharyngeal malformation19. On the basis of these findings, we proposed that TGFβ–SMAD2/3 signalling may contribute to organizer formation and/or organizer-mediated induction in M. leidyi. All identified receptors of the TGFβ family in M. leidyi share the canonical receptor architecture: an extracellular domain, a single-pass transmembrane segment and an intracellular serine–threonine kinase domain. Moreover, the three type I receptors (encoded by MlTgfrIa–MlTgfrIc) contain a glycine–serine repeat region immediately adjacent to the kinase domain, a characteristic consistent with their classification as type I (ALK) receptors20. Phylogenetic analyses do not clearly separate these receptors into BMP, Activin or Nodal subclasses. Therefore, we refer to them here simply as TGFβ type I and type II receptors19. Accordingly, to assess the role of TGFβ type I receptor activity in M. leidyi organizer formation and oral structure development, we treated embryos with two independent TGFβ receptor type I inhibitors, A83-01 and SB431542. Inhibition of TGFβ–SMAD2/3 signalling led to pharyngeal malformations19 (Extended Data Fig. 5a–i) and the downregulation of MlBrachyury and another organizer-associated gene, MlLhx1/5, in the developing mouth of M. leidyi. By contrast, expression of the aboral marker MlLhx3/4 remained unchanged (Extended Data Fig. 5j–l). In vertebrates, the Lhx1/5 homologue LHX1 (also known as LIM1) functions as an organizer-associated gene for which expression depends on TGFβ signalling mediated by SMAD2 and SMAD3 (TGFβ–SMAD2/3 signalling), and is specifically activated by Nodal and/or Activin ligands21,22. By contrast, in bilaterians, Brachyury expression typically depends on both TGFβ–SMAD2/3 and β-catenin signalling23,24. This characteristic is consistent with the knowledge that coordinated β-catenin–TGFβ–SMAD2/3 regulatory interactions have deep evolutionary roots and were already evident in pre-bilaterian lineages such as cnidarians (for example, Hydra)