A prognostic human brain network for diffuse midline glioma
Abstract
Diffuse midline gliomas (DMGs) are near-universally lethal tumours of the childhood central nervous system1,2. In animal models, DMGs form brain-wide integrated networks through neuron-to-glioma synapses3,4,5,6 and glioma-to-glioma gap junctional coupling3. This extensive connectivity robustly promotes the growth and invasion of DMG3,4,5,6,7,8,9 and other glial malignancies10,11,12 through paracrine mechanisms and direct neuron-to-glioma synapses. However, the organization and clinical implications of these connections in the living human brain remain to be elucidated. Here, we develop tumour network mapping to compute the brain-wide connectivity profile of DMG, defining a conserved brain network across pontine and thalamic DMG associated with patient short-term survival (DMG network). Tumour functional connectivity with the DMG network was independently predictive of patient overall survival across two external validation cohorts. Tumour growth mapped to DMG network-specific trajectories and peak in-network neurometabolic changes across development spatiotemporally aligned with the peak age incidence of DMG. Analyses of single-nucleus RNA sequencing data confirmed diverse synaptic gene enrichment in high-connectivity DMG. Strikingly, incidental surgical resection of high-connectivity thalamic DMG tissue conferred a significant survival advantage. Collectively, these data define a conserved and prognostically important brain network in children with DMG, consistent with the hypothesis that DMGs exploit otherwise healthy brain circuits to promote tumour growth.
Similar content being viewed by others
Pharmaco-proteogenomic profiling of pediatric diffuse midline glioma to inform future treatment strategies
Microglia in diffuse midline glioma contribute to extracellular matrix remodelling and cancer cell invasion

Spatial transcriptomics reveals niche-specific enrichment and vulnerabilities of radial glial stem-like cells in malignant gliomas
Main
Diffuse intrinsic pontine glioma (DIPG) and other diffuse midline gliomas (DMGs) are the leading cause of solid-tumour-related death in children which despite clinical trials over the past two decades, retain notorious therapeutic resistance and near-universal lethality2,13. A major clinical challenge limiting the efficacy of conventional therapeutic approaches, and recognized in original definitions of the disease, is the extensive infiltration of healthy brain parenchyma14. This distributed, brain-wide pattern of tumour progression, including to distant brain regions such as the frontal and temporal poles, is insufficiently explained by current models of tumour evolution15,16,17. Recent work has established DMG integration and communication with otherwise healthy neural circuits through both paracrine signalling (brain-derived neurotrophic factor (BDNF) and neuroligin-3 (NLGN3))4,7,8 and bona fide, electrophysiologically functional neuron-to-glioma synapses across a diverse neurotransmitter repertoire, including glutamatergic, calcium-permeable AMPA receptor-mediated3,4; cholinergic M1–M3 receptor-mediated5; and GABAergic GABAA receptor-mediated6 neuron-to-glioma synapses. In animal models, depolarization of glioma cell membranes drives tumour growth through voltage-dependent mechanisms that remain to be fully elucidated3,4. Neuronal cell somata local to or distant from and projecting onto malignant cells engage brain-wide neural populations in glioma circuits, activity from which acts as a putative driver of tumour progression5,9,18. For example, long-range cholinergic projections from the midbrain pedunculopontine and laterodorsal tegmental nuclei, respectively, promote the circuit-specific growth of pontine and thalamic DMG in preclinical models5. Although the impact of other neuromodulatory neurons remains to be fully explored, early preclinical reports suggest that serotonergic projections from the dorsal and median raphe promote the circuit-specific growth of pontine, thalamic and cortical DMG, while noradrenergic projections from the locus coeruleus also promote DMG growth9,18.
Thus far, attempts to map the brain-wide neural circuits implicated in tumour growth have been restricted to mouse models, which offer an incomplete representation of human-specific neurobiology and therapeutic relevance. Further, the distinct spatiotemporal pattern of DMG incidence, such that pontine and thalamic tumours peak in incidence during early childhood and early adolescence, respectively, suggests tumour exploitation of an underlying neurodevelopmental process. Here, using patient clinical data and human paediatric connectomic data, we define the spatial topography of brain network connections associated with short-term survival in children with DMG and delineate circuit-specific trajectories of DMG tumour growth.
Tumour network mapping
We first studied a discovery cohort of 125 children aged less than 18 years with primary pontine DMG/DIPG (n = 106), or thalamic DMG (n = 19), treated at Great Ormond Street Hospital for Children (GOSH; Fig. 1a and Supplementary Table 1). As pontine biopsies were historically not performed19, children meeting clinical diagnostic criteria for DIPG20,21,22 and with a disease course consistent with short-term (<18-month)23 overall survival were included. In line with the World Health Organisation 2021 definition of the disease1, biopsied tumours are classified as DMG, H3K27-altered, while non-biopsied pontine tumours are classified as DIPG. Throughout, reproducibility of findings is demonstrated for the combined (DMG/DIPG) patient cohort and across restricted cohorts of children with biopsied pontine or thalamic DMG, H3K27-altered. Patient tumours were segmented on preoperative brain magnetic resonance imaging (MRI) and mapped to a standard paediatric template (Extended Data Fig. 1a–c). Tumour locations were compared between patients with short- and long-term overall survival using voxel-based lesion-symptom mapping (VLSM). Univariate VLSM across discovery cohort pontine and thalamic tumours identified a significant association between patient short-term survival and tumour pontine location (Extended Data Fig. 1d,e), reflecting the known shorter overall survival of children with pontine versus thalamic DMG2. Aiming to identify specific voxels within the pons or thalamus associated with patient short- versus long-term survival, we performed separate univariate VLSM for pontine and thalamic tumour locations and multivariate VLSM across all tumour locations. Across both methods, no voxels associated with short-term survival were identified (Fig. 1b and Extended Data Fig. 1f–h). To test for any prognostic impact of subthreshold VLSM results (defined as weak, non-significant voxelwise associations with short-term survival), we collected data from an independent cohort of children with primary pontine DMG/DIPG (n = 80) or thalamic DMG (n = 45) treated at one of three international centres or included in the HERBY trial (NCT0139094824; Supplementary Table 1), with identical inclusion criteria. The weighted overlap of each tumour in this external validation cohort with univariate and multivariate VLSM maps, respectively, was computed and averaged to generate an overlap score25. No survival difference was identified between patient tumours stratified into high- and low-overlap risk groups using a median cut-off across univariate and multivariate VLSM (Extended Data Fig. 1i–l). Given this lack of association between tumour location and patient survival, the influence of neural activity on DMG growth in preclinical models3,4,5,6,7,8 and the fact that human DMG/DIPG exhibits a common pattern of brain infiltration (Extended Data Fig. 2), we hypothesized that tumour connectivity with an underlying brain network would better explain the observed variance in patient survival.
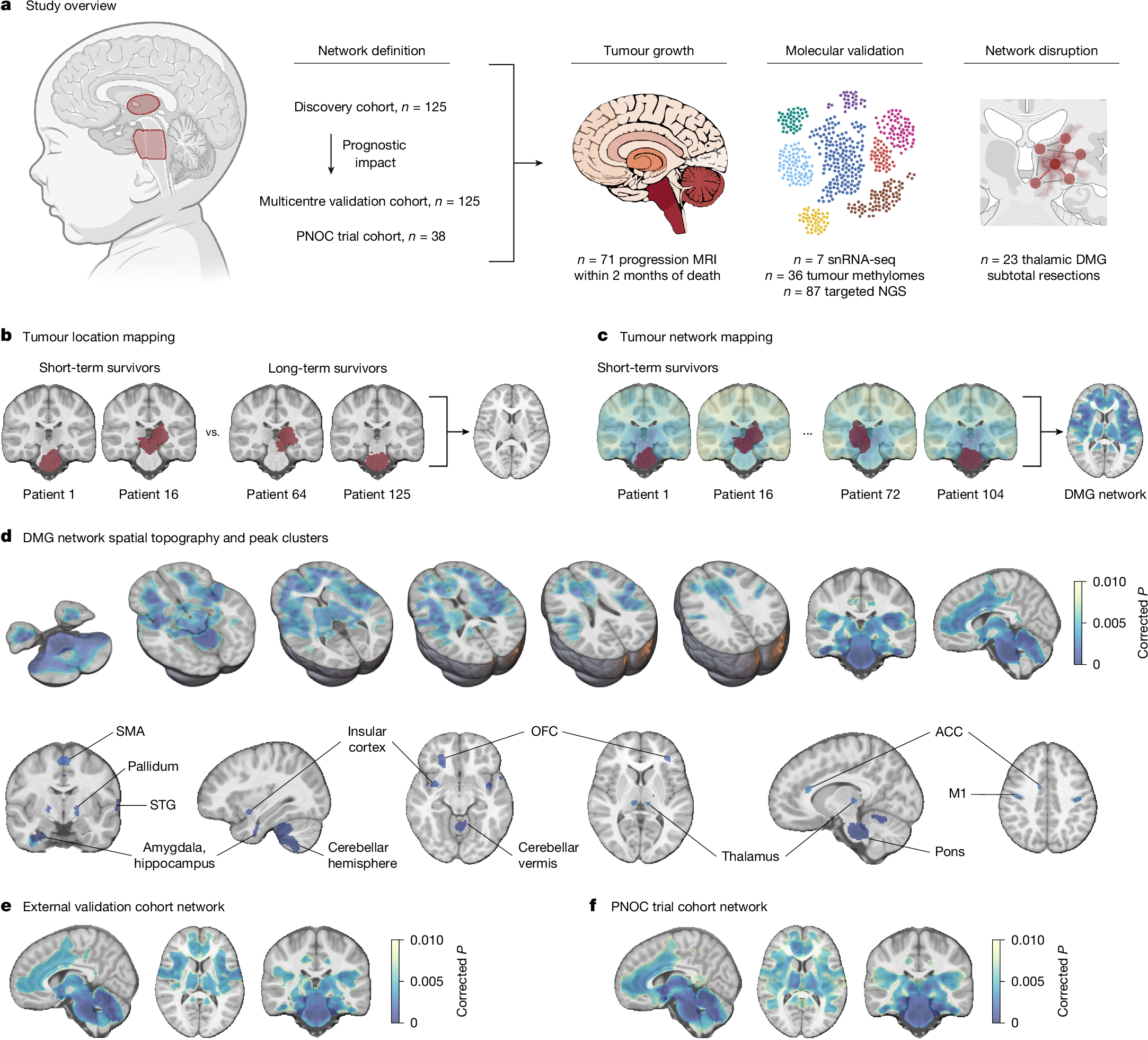
a, Overview of the study methodology. NGS, next-generation sequencing. b, The tumour locations of short-term and long-term survivors of pontine DMG/DIPG and thalamic DMG from the discovery cohort (n = 125; left, red) were compared using VLSM; however, tumour locations did not explain the observed variance in patient overall survival (right; PFWE > 0.05). c, Tumour network mapping was performed to compute the network of brain regions connected to each patient’s tumour location using resting-state functional connectivity data from 1,000 healthy children aged 9.0 ± 0.2 years (the human connectome). Tumour network maps across patients with short-term overall survival (n = 106) were statistically compared to identify brain regions that are significantly associated with shorter patient overall survival (right; network peaks are shown in dark blue), hereafter termed the DMG network. d, DMGs exhibit a specific pattern of brain functional connectivity associated with patient short-term overall survival. This includes strong positive connectivity with the pons, thalamus (centromedian, medial pulvinar and ventral intermediate nuclei), motor cortex (M1 and supplementary motor area (SMA)), orbitofrontal (OFC) and insular cortices, limbic system (amygdala, hippocampus, basal ganglia and anterior cingulate cortex (ACC)), superior temporal gyrus (STG) and cerebellum (lobules I–V, VIII and X). e,f, Identical tumour network mapping performed across two independent external validation cohorts (external validation cohort network (e) and PNOC trial cohort network (f)) reproduced DMG network topography. n = 125 children with primary pontine DMG/DIPG and thalamic DMG, n = 108 short-term survivors (e); n = 38 children with biopsy-confirmed pontine DMG, n = 29 short-term survivors (f). For e, DMG network whole brain spatial ρ = 0.952, label permutation P < 0.001; spin spatial ρ = 0.974, spin permutation P < 0.001. For f, DMG network whole brain spatial ρ = 0.911, label permutation P < 0.001, spin spatial ρ = 0.885, spin permutation P < 0.001. Statistical maps were generated using threshold-free cluster enhancement and corrected for multiple comparisons using familywise error (FWE) rate correction for multiple testing. Two-tailed PFWE < 0.01 was considered significant for connectivity data. Brain slices are shown in radiological orientation. The diagram in a was created using BioRender; Sidpra, J. https://BioRender.com/vi1xgg0 (2026).
Lesion network mapping is a computational neuroimaging method that harnesses population-level (n = 1,000) connectomic data to model common brain network connections across different lesion locations26,27,28. It has successfully identified brain networks and therapeutic targets across neurological and psychiatric disorders, including epilepsy29,30, addiction31 and depression32. Here, we develop tumour network mapping, using a lesion network mapping framework, to identify common brain network connections associated with the short-term survival of patients with DMG, irrespective of tumour primary pontine or thalamic location. Tumour volume was included as a covariate in tumour network mapping analyses to account for potential confounding by lesion size26,29,31. In this cohort, tumour volume was not associated with overall survival (univariate Cox hazard ratio (HR) per 1,000 mm3, increase of 1.00; 95% confidence interval (CI) = 0.99–1.01; P = 0.569), consistent with previous clinical reports33,34. Given our aim to identify polysynaptic and direct structural connections between brain regions, whole-brain-to-tumour connectivity analyses were performed across paediatric resting-state functional connectivity (functional MRI, fMRI)35 and structural connectivity (diffusion-weighted MRI, dMRI)36,37 data. These analyses identified a distributed brain network associated with shorter patient overall survival (Fig. 1c). More specifically, tumours exhibiting greater positive functional connectivity with the pons, thalamus, motor cortex, insular and orbitofrontal cortices, limbic system (amygdala, hippocampus, basal ganglia and cingulate cortex), superior temporal gyrus and cerebellum were associated with shorter patient overall survival (Fig. 1d and Supplementary Table