Structure of the pre-initiation complex explains CMGE biogenesis
Abstract
When cells enter S phase, bidirectional DNA replication is initiated through the kinase-regulated recruitment of three activators (Cdc45, GINS and Pol ε) to a duplex-DNA-loaded double hexamer of minichromosome maintenance (MCM) ATPases. Together, these proteins form two CMGE helicases that establish divergent replication forks as they become separated1. Here, to gain an understanding of CMGE biogenesis, we reconstituted the pre-initiation complex with purified yeast proteins. The cryo-electron-microscopy structure shows a set of firing factors caught in the act of assembling two symmetrical CMGEs. We show how stepwise complex formation reshapes MCM in preparation for DNA opening, and we explain how ATP promotes firing-factor ejection and CMGE maturation. We find that although Sld2 facilitates the recruitment of GINS to MCM, as expected, it also aids the efficient separation of the CMGE dimer, and is essential for the ejection of the lagging strand from MCM. These findings have direct implications for our understanding of the metazoan Sld2 orthologue, RECQL4, and point to a replication-fork establishment mechanism that is conserved across eukaryotes.
Main
DNA replication must occur only once per cell cycle to maintain genome stability. To achieve this, eukaryotes have evolved to temporally separate the loading of the replicative helicase from its activation1. During the G1 phase of the cell cycle, two copies of the MCM motor of the replicative helicase are loaded as an inactive double hexamer (DH) onto DNA replication origins2,3. Activation occurs after S-phase transition and involves the recruitment of Cdc45 and the tetrameric GINS (Go-Ichi-Ni-San) complex to MCM, together forming the Cdc45–MCM–GINS (CMG) helicase4,5. CMG assembly occurs under the control of three kinases. It is promoted by the Dbf4-dependent kinase (DDK)6,7,8 and Cdc28–Clb5 (hereafter, CDK), the activity of which increases when cells enter S phase9,10,11. CMG assembly is inhibited by the checkpoint kinase Rad53, which blocks late origin firing if DNA damage is detected9,10,11,12,13. DDK selectively phosphorylates DNA-loaded DHs14. A heterodimeric firing factor composed of Sld3 (essential) and Sld7 (dispensable) then recognizes the phosphorylated DH and recruits Cdc45 to MCM (refs. 10,11,15). An N-terminal truncation of Mcm4 bypasses the requirement for DDK in cells7,8 and cryo-electron microscopy (cryo-EM) work has shown that phosphorylation causes N-terminal Mcm4 to become unstructured16,17. Whether Sld3 engages an epitope that is unmasked after N-terminal Mcm4 phosphorylation or whether it merely reads phosphorylated (phospho-) Mcm4 sites remains to be established. Also, although we know that Rad53 prevents CMG formation by targeting Sld3 (alongside DDK)13, the mechanism is unclear. The second activating kinase, CDK, targets two firing factors, Sld3 and Sld2. The latter was previously implicated in GINS recruitment. Phospho-Sld2 and phospho-Sld3 are recognized by the Dpb11 ‘phospho-reader’10,11. The leading-strand polymerase Pol ε (formed of Pol2, Dpb2, Dpb3 and Dpb4) also contributes to CMG assembly and becomes part of the holohelicase, CMGE. In particular, the N-terminal domain of Dpb2 supports CMG formation in cells18, and a complex containing Dpb2 and the C-terminal half of Pol2 achieves GINS recruitment and replication initiation in a reaction reconstituted with purified yeast proteins19.
In the inactive DH, an Mcm7-specific N-terminal insertion (NTI) from one MCM hexamer reaches across the DH interface and protects the N-terminal A domain of Mcm5 of the opposing hexamer20 (Fig. 1a). The same Mcm5 A-domain site is engaged by GINS in the CMG, implying that Mcm7 must let go of Mcm5 for GINS to bind21,22. How this happens is unknown. Likewise, it is established that Cdc45 and GINS recruitment occur sequentially23,24,25. Whether recruitment of each component involves only one MCM hexamer or both MCM hexamers at once is debated25,26,27.
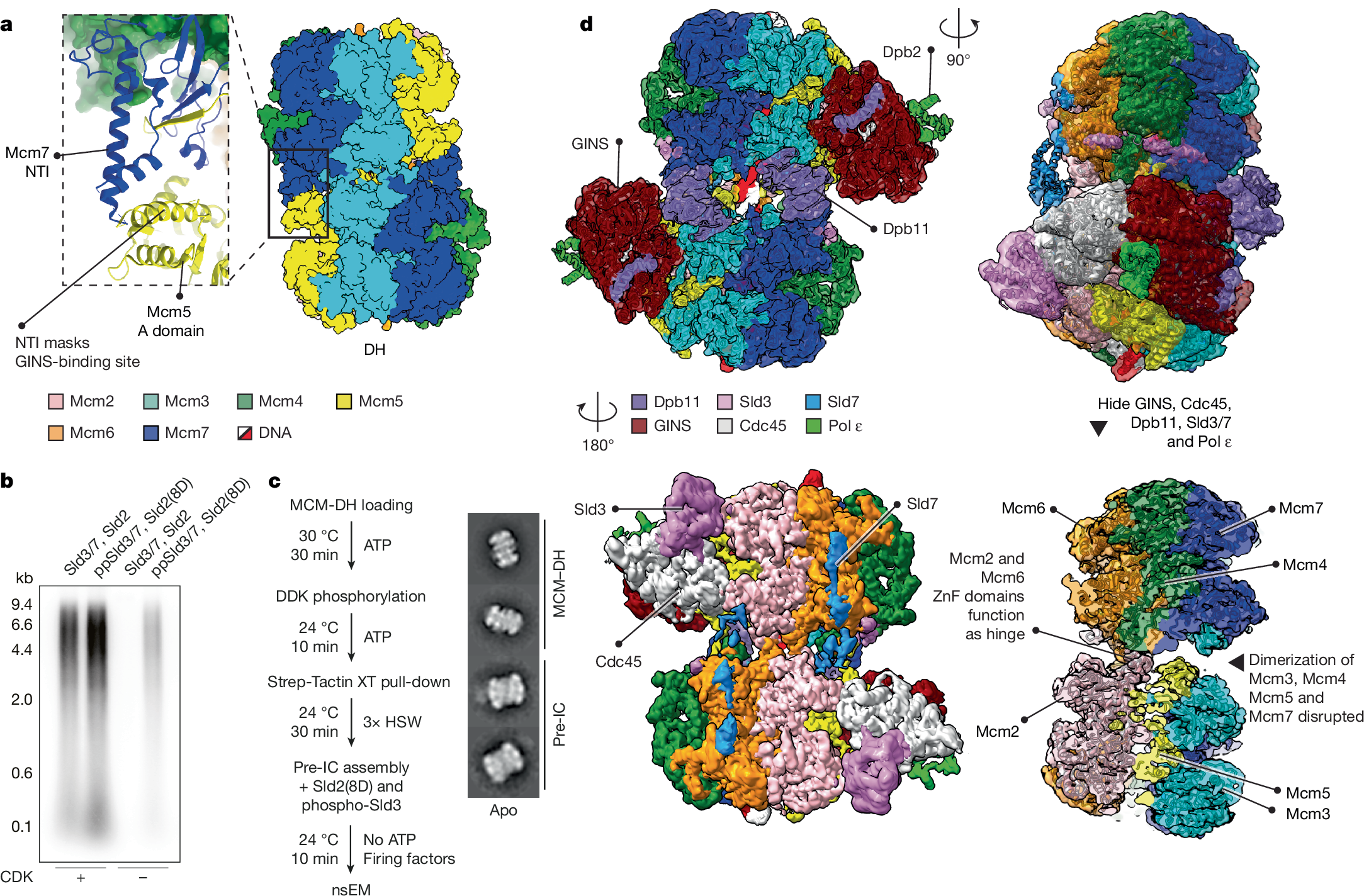
a, In the double hexamer (DH; Protein Data Bank (PDB) ID: 7P30), the Mcm7 NTI element from one MCM ring protects the N-terminal Mcm5 A domain of the opposed ring. b, CDK-prephosphorylated Sld3/7 (ppSld3/7) and the Sld2(8D) phosphomimetic mutant support origin-dependent replication reconstituted in a test-tube, albeit at reduced levels (experiment performed twice). The low levels could be due either to the Sld2(8D) phosphomimetic mutations covering only a subset of CDK sites, or to inefficient phosphorylation of Sld3 in the isolated prephosphorylation reaction, compared with the complete initiation mix. For gel source data, see Supplementary Fig. 1. c, CMG assembly in the absence of ATP, using ppSld3/7 and Sld2(8D), enables formation of the pre-IC. HSW, high-salt wash. Right, representative 2D classes. d, Cryo-EM structure of the pre-IC. The two MCMs are split on one side, with Mcm2 and Mcm6 functioning as a hinge. ZnF, zinc finger domain.
Origin activation is controlled by ATP binding and hydrolysis. MCM loading, for example, requires ATP hydrolysis28,29, so that, by the time a DH is formed, eight of its twelve subunits are bound to ADP (ref. 14). Firing-factor recruitment to the DH promotes ADP release, and ATP binding by MCM achieves stable double CMG–Pol ε (ATP–dCMGE) complex formation, which nucleates DNA melting24,30. It has been postulated that a pre-initiation complex (pre-IC) exists31, in which Sld2, Sld3–Sld7 (hereafter, Sld3/7) and Dpb11 all bind to the DH at the same time, while also recruiting Cdc45, GINS and Pol ε (refs. 24,32). However, the pre-IC has not so far been isolated, and we do not know how Sld2, Sld3/7 and Dpb11 are ejected to achieve ATP–dCMGE maturation24,30. Finally, whether the role of Sld2 is conserved across eukaryotes is unclear. In fact, Sld2 functions in CMGE assembly in yeast, whereas the metazoan orthologue, RecQL4, is involved in a loosely defined downstream activation step. To address these issues and understand replisome biogenesis, we took a biochemical reconstitution approach, combined with cryo-EM imaging and single-particle reconstruction.
Cryo-EM structure of the pre-IC
In previous cryo-EM work, we established that by the time the ATP–dCMGE complex is formed at an origin, all of the factors involved in its assembly process have been released30. We reasoned that, if ATP binding by MCM promotes this release, forming dCMGE in a buffer that lacks ATP might mean that the firing factors Sld2, Sld3/7 and Dpb11 are retained at origins, allowing us to reconstitute the pre-IC. Obtaining enough sample for cryo-EM analysis in such conditions is a challenge. Moreover, ATP is required for DH phosphorylation by DDK (ref. 33), as well as Sld2 and Sld3 phosphorylation by CDK (refs. 10,11). To circumvent this issue, we isolated the phospho-DH on origin DNA30, in a buffer lacking ATP (Supplementary Fig. 2a–e). Inspired by previous work24, we also phosphorylated recombinant Sld3/7 using CDK and repurified it in ATP-free conditions (prephosphorylated Sld3/7; hereafter, ppSld3/7; Supplementary Fig. 2f). We also cloned, expressed and purified a phosphomimetic variant of Sld2 containing 8 of 11 aspartate substitutions (hereafter, Sld2(8D)), previously shown to support Dpb11 binding and CDK bypass in cells10,34 (Supplementary Fig. 2g). We then tested ppSld3/7 and Sld2(8D) in a DNA replication reaction reconstituted in vitro with purified proteins19. DNA replication could be established in the absence of CDK with these reagents, although at reduced levels, compared with reactions containing CDK (Fig. 1b). Despite the reduction, ATP–dCMGE complexes30 could be assembled efficiently using ARS1-loaded phospho-DH, ppSld3/7 and Sld2(8D) in the absence of CDK, as observed by negative-stain electron microscopy (nsEM; Supplementary Fig. 3a,b). We then asked whether GINS and Cdc45 could be recruited to MCM using ppSld3/7 and Sld2(8D), in the absence of CDK and any nucleotide. nsEM two-dimensional (2D) averaging revealed a complex reminiscent of ATP–dCMGE30, but with MCMs engaged in tighter interaction and with no recognizable Pol ε density—at least at the limited resolution achieved with negative staining (Fig. 1c). Unlike ATP–dCMGE, the new ATP-free complex became disassembled when purified in high-salt conditions (Supplementary Fig. 3c–e). This observation agrees with previous western blot evidence that high-salt-resistant association of GINS and Cdc45 with licensed-origin DNA requires ATP binding24. To determine the composition of the ATP-free high-salt-sensitive assembly, we solved the cryo-EM structure (Extended Data Figs. 1 and 2). Inspection of the resulting density map, refined to 3.4-Å resolution after symmetry expansion35 (or 3.2 Å for the locally refined asymmetric unit), revealed a quasi-symmetric (flexible) assembly of two CMGs (Extended Data Table 1). Compared with the DH, Mcms 5, 3, 7 and 4 are disengaged across the two rings. Homodimerization through Mcm2 and Mcm6 persists as observed20 in the DH, with zinc finger domains acting as a hinge. Unoccupied density could be assigned to two copies of Sld3, Sld7, Dpb11 and the Dpb2 subunit of Pol ε, which, together with two CMG assemblies, form the pre-IC complex (Fig. 1d).
Kinase-regulated Mcm4 engagement by Sld3
Previous studies16,17,36 have established that docking of DDK onto the DH dislodges the N-terminal tail of Mcm4 from the A domain of Mcm4 (A4; Fig. 2a,b). A4 remains occupied by the Dbf4 subunit of DDK, while the Cdc7 kinase subunit phosphorylates the N-terminal Mcm4 tail that has become solvent-exposed (Fig. 2a,c). When DDK releases the DH, the negatively charged phospho-Mcm4 tail does not return to binding the electronegative A4 site, probably owing to electrostatic repulsion16,17 (Fig. 2a,d). Within the pre-IC, we find that the newly exposed A4 epitope becomes engaged by a C-terminal Sld3 α-helix, which in turn extends towards the two neighbouring subunits on either side of Mcm4. This same interaction was also predicted using AlphaFold 337. N-terminally, Sld3 touches the Mcm6 A domain and C-terminally, it engages the Mcm7 NTI element (Fig. 2e). This Sld3 interaction might contribute to dislodging Mcm7 from its position across the two hexamers, where it protected the Mcm5 A domain in trans. We expressed and purified a truncated Sld3/7 complex lacking the Mcm4-binding site of Sld3 (∆515–538), to test whether this variant can still form the pre-IC and support DNA replication in reconstituted reactions. We found that, compared with the wild-type protein, both functions were almost completely blocked when using the truncated variant (Fig. 2f,g). Our results agree with previous biochemical data, which identified the same Sld3 region as being essential for MCM binding23. In addition to the interaction with Mcm4 described above, Sld3 has been observed to bind specific phosphorylated segments of Mcm4 and Mcm6 independently (not observed in our structure). This indicates that Sld3 has a distinct phospho-reader role, which complements its function in recognizing a DDK-dependent structural change in MCM (ref. 23).
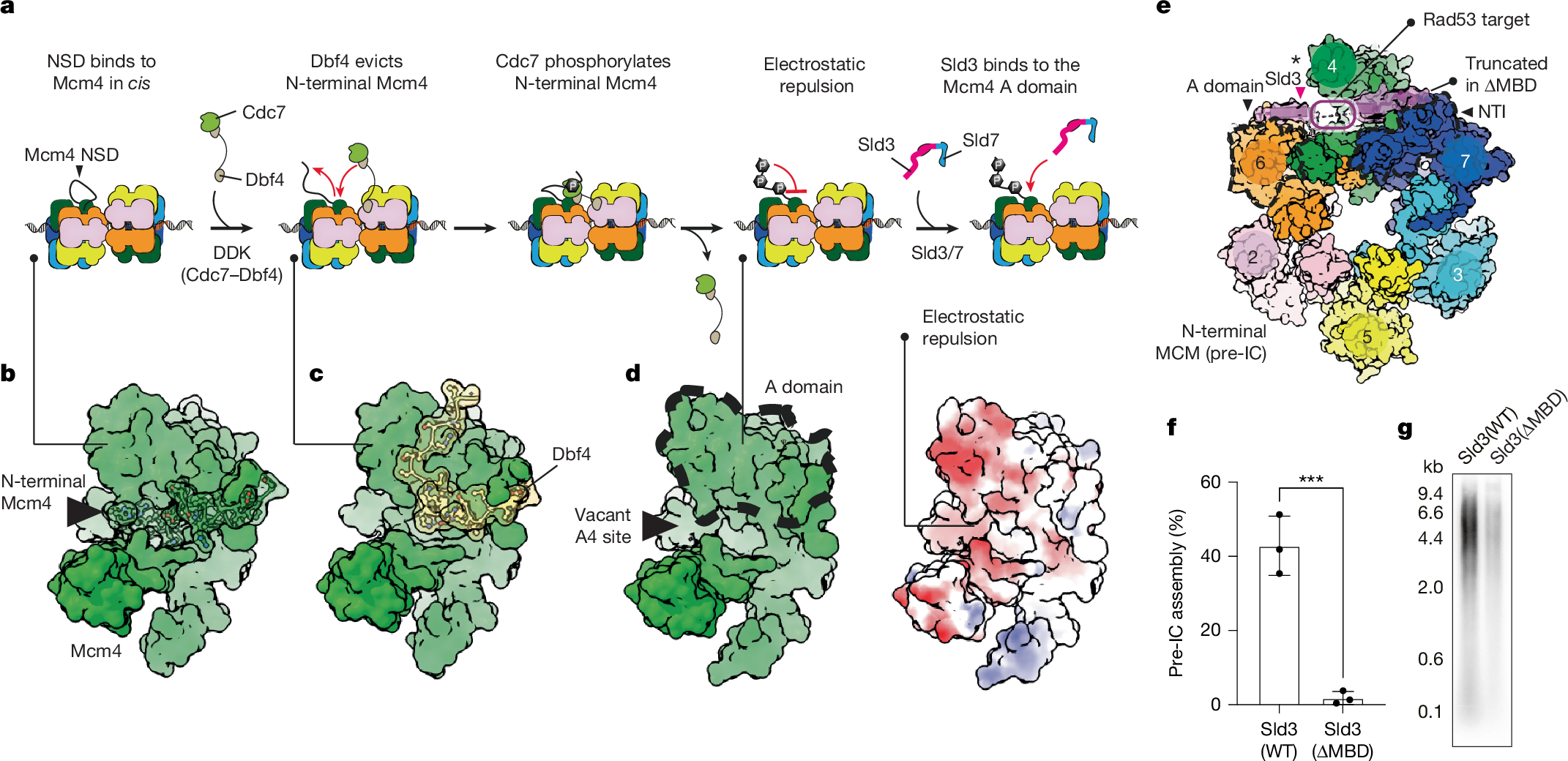
a, Cartoon of the sequence of events leading to Sld3 recruitment. First, the DDK subunit Dbf4 dislodges N-terminal Mcm4 from the Mcm4 A domain. Then, DDK phosphorylation of N-terminal Mcm4 prevents its re-engagement with the A domain. Finally, Sld3 binds to the newly exposed Mcm4 A-domain epitope (vacant A4 site). b, Structure of non-phosphorylated Mcm4 (PDB: 7V3U). c, Structure of Mcm4 engaged by the Dbf4 subunit of DDK (PDB: 7PT7). d, Structure of phosphorylated Mcm4 (PDB: 7P30). Right, surface representation of phosphorylated Mcm4, coloured by electrostatic potential. e, Sld3 binds to the Mcm4 A domain as well as to the neighbouring NTI Mcm7 element and Mcm6 A domain. MBD, MCM-binding domain. The asterisk indicates Sld3 binding to the Mcm4 A domain. f,g, Truncation of the Sld3-MBD impairs pre-IC assembly (f; ***P = 0.0010 by two-tailed t-test; mean ± s.d.; experiment performed three times) and replication reconstituted in a test-tube (g; experiment performed twice). WT, wild type. For gel source data, see Supplementary Fig. 1.
Rad53 halts origin firing by targeting Sld3, as well as the Dbf4 subunit of DDK. In the Supplementary Results, we show that Rad53 phosphorylates a segment of Sld3 that is involved in binding to the A4 site, hence blocking origin firing by preventing Sld3 recruitment (Supplementary Figs. 4 and