Coral microbiomes as reservoirs of unknown genomic and biosynthetic diversity
TL;DR
Coral reef microbiomes, especially from reef-building corals, contain vast unknown genomic and biosynthetic diversity, with over 90% of microbial species lacking prior genomic data. Their biosynthetic potential rivals traditional sources like sponges, highlighting the need for conservation to preserve these molecular resources.
Key Takeaways
- •Coral microbiomes host over 4,000 microbial species, with 90% previously uncharacterized genomically, indicating a vast reservoir of unknown diversity.
- •The biosynthetic potential of reef-building coral microbiomes is as rich or richer than that of sponges, a traditional source of bioactive compounds.
- •Microbial species in coral microbiomes are highly host-specific, with minimal overlap between coral types and seawater, emphasizing unique ecological niches.
- •New groups of Acidobacteriota in coral microbiomes encode unknown enzymology, offering promising avenues for biotechnology and protein engineering.
- •Conservation of coral reefs is crucial to prevent loss of this molecular diversity, given their decline and role as vital reservoirs for bioactive metabolites.
Tags
Abstract
Coral reefs are marine biodiversity hotspots that provide a wide range of ecosystem services1. They are reservoirs of bioactive metabolites, many produced by microorganisms associated with reef invertebrate hosts2. However, for the keystone species of coral reefs—the reef-building corals—we still lack a systematic assessment of their microbially encoded biosynthetic potential and the molecular resources at stake due to the alarming decline in reef biodiversity. Here we analysed microbial genomes reconstructed from 820 reef-building coral samples of three representative coral genera collected at 99 reefs across 32 islands throughout the Pacific Ocean (Tara Pacific expedition)3. By contextualizing our analyses with the microbiomes of other reef species, we found that only 10% of the 4,224 microbial species and less than 1% of the 645 species exclusively identified in Tara Pacific samples had genomic information available. Furthermore, the biosynthetic potential of reef-building coral microbiomes rivalled or surpassed that of traditional natural product sources such as sponges. Among the biosynthetically rich bacteria in the reef microbiome, we identified new groups of Acidobacteriota that encode previously unknown enzymology, in turn opening promising avenues for functional protein engineering. Together, this study underscores the importance of conserving coral reefs as vital reservoirs of molecular diversity.
Similar content being viewed by others
Diversity of the Pacific Ocean coral reef microbiome

The coral microbiome in sickness, in health and in a changing world
Integrative omics framework for characterization of coral reef ecosystems from the Tara Pacific expedition
Main
Coral reefs are one of the most biodiverse and productive ecosystems on Earth. Despite covering less than 0.2% of the ocean floor, they are home to a third of all named marine multi-cellular species4,5. Coral reefs provide a wide range of ecosystem services, such as food, livelihoods and coastal protection, to millions of people around the globe, and they serve as a source of bioactive metabolites1,2. However, climate change, emerging diseases and other anthropogenic stressors have caused a decline in live coral cover of more than 50% since the 1950s1,6. Given the projections of further reef decline7, there is a pressing need to capture what is at stake under this continued biodiversity loss.
The biodiversity, productivity and structural complexity of reef systems are linked fundamentally to the ecological functions provided by calcareous skeleton-forming (reef-building) corals, such as stony and fire corals1. Like other organisms, these sessile invertebrates depend on a diverse community of microorganisms (microbiome)8,9. The microbiome of corals provides its host with vital nutrients, such as carbon, nitrogen and phosphorus, as well as vitamins and essential amino acids10. Furthermore, it supports its host in coping with changing environmental conditions11 and can protect it from infectious diseases12. Microbes associated with reef-building corals are suggested to produce bioactive metabolites to fend off pathogens, predators and competitors12. However, although diverse metabolites such as anti-microbial13, anti-inflammatory14 and anti-tumour15 agents have been discovered (with some undergoing clinical trials16) in other reef invertebrates2 such as sponges and soft corals, little is known about the bioactive potential of reef-building coral microbiomes.
Historically, the discovery of bioactive metabolites has relied on screening chemical extracts from either the invertebrate host, which may depend on the supply of unsustainable amounts of animal biomass17, or from microbial producers isolated from their hosts15. The latter approach is, however, constrained by our limited ability to cultivate most microorganisms under standard laboratory conditions18. Furthermore, both methods are prone to the persistent challenge of rediscovering the same or similar metabolites19. As a more recent strategy, metabolic pathways linked to biosynthetic gene clusters (BGCs) that encode the synthesis of bioactive compounds can be discovered by screening reconstructed genome sequences20. These genomes may originate from microbial culture collections18 as well as from uncultivated single cells or whole microbial communities (metagenomes) from, in principle, any environment or host organism21,22,23,24. However, for the microbiome of reef-building corals, such genomic information remains scarce18,25.
We thus aimed to explore systematically the genome-resolved diversity, host-specificity and BGC-encoded biosynthetic potential of reef-building coral microbiomes, compare them with the microbiomes of other reef hosts (such as sponges) and the surrounding environment, and determine whether corals host any BGC-rich lineages as promising biotechnological targets. To this end, we reconstructed more than 13,000 metagenome-assembled genomes (MAGs) from reef-building coral samples collected as part of the Tara Pacific expedition3 (Supplementary Table 1) and from publicly available coral reef metagenomic datasets (Supplementary Table 2). For almost 90% of the 4,224 microbial species in total, or more than 99% of those from Tara Pacific samples for which we reconstructed MAGs, no genome-resolved information was available previously. Coral and sponge microbiomes were largely host-specific and we found the biosynthetic potential (per microbial species) in reef-building corals (particularly fire corals) to be as rich as, or even richer than, that of sponges or the surrounding waters. By detecting new, biosynthetically rich bacterial lineages and characterizing unusual enzymology and bioactive compounds within coral-associated Acidobacteriota26 spp., our work clearly underscores not only the value of reef-building corals from a biotechnological perspective, but also the implications of their potential loss.
Coral reef microbiome genomic resources
To fill the gap in the availability of microbial genome data from reef-building corals, we collected 820 metagenomes from two stony coral genera (Porites and Pocillopora) as well as fire corals (Millepora) around 32 islands (99 reefs) throughout the Pacific Ocean as part of the Tara Pacific expedition3 from 2016 to 2018 (Fig. 1a,b; Supplementary Table 1; for details on coral host lineages, see ref. 27). To facilitate a comprehensive assessment of the phylogenomic novelty and biosynthetic potential of reef-building coral microbiomes, and to contextualize this information across different coral reef-inhabiting species (such as sponges and soft corals), we supplemented the dataset with existing metagenomes from 412 coral samples (from 29 genera, including 22 stony and five soft coral genera, as well as from black and fire corals) and 371 sponge samples (from 32 genera) (Fig. 1c,d; Extended Data Fig. 1a and Supplementary Table 2).
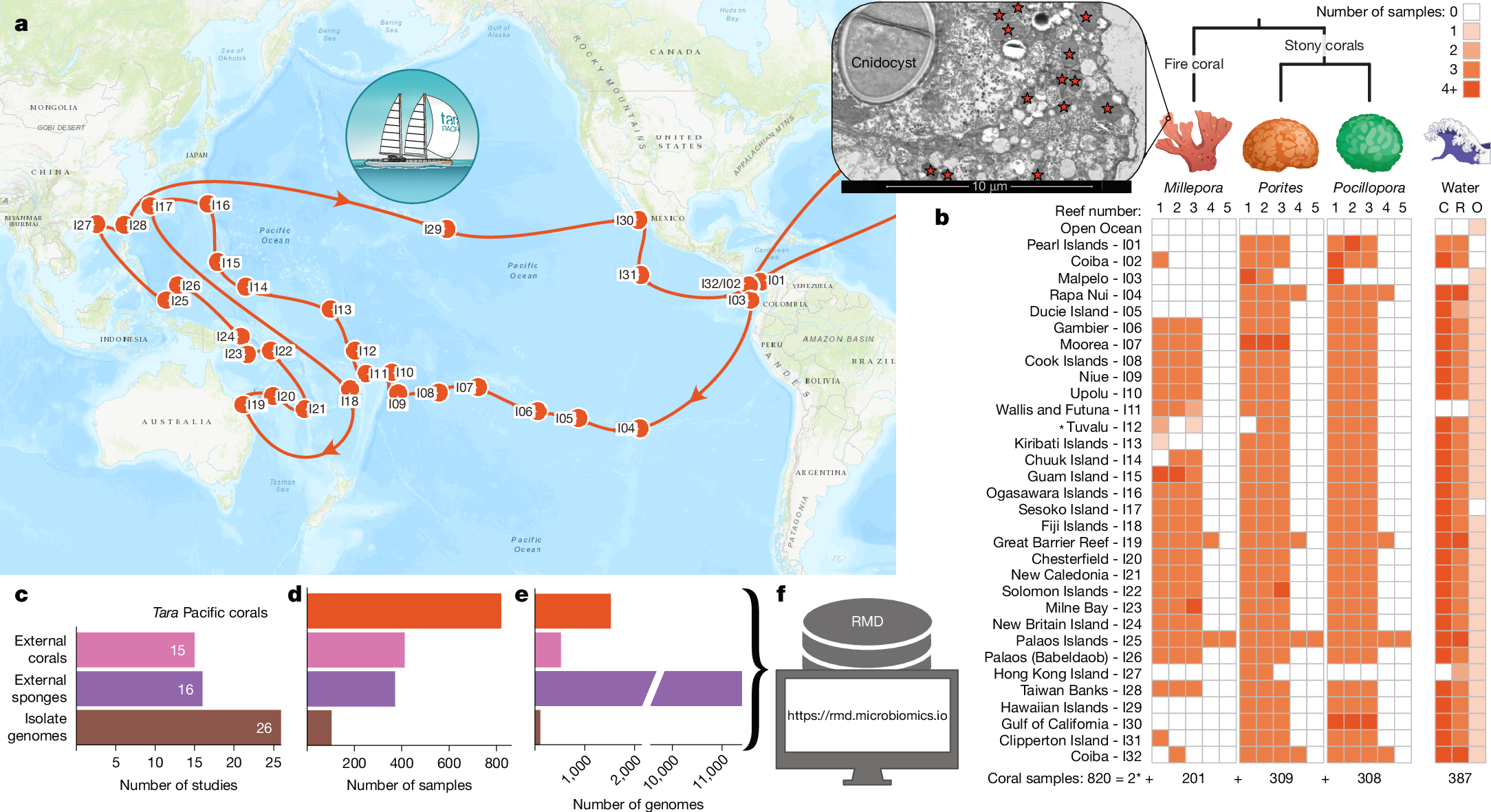
a, The Tara Pacific expedition (2016–2018)3 included the sampling of corals at 99 reefs across 32 islands throughout the Pacific Ocean. b, At each reef, Millepora, Porites and Pocillopora colonies (asterisk, plus (exceptionally) two Heliopora specimens at one reef in Tuvalu) were sampled28, resulting in a total of 820 reef-building coral-associated metagenomes (Supplementary Table 1). In addition, the plankton microbiome was collected from the water surrounding Pocillopora colonies, from representative water within each reef, as well as from oceanic water28, resulting in 387 metagenomes (Supplementary Table 1). C, coral-surrounding water; R, reef water; O, open ocean water. Inset, TEM image of a Millepora tissue sample, with bacteria-sized cells in the ectoderm indicated by stars (Extended Data Fig. 1c; Methods). c,d, To contextualize the data generated from the Tara Pacific expedition, we aggregated a total of 412 coral (from 29 genera) and 371 sponge (from 32 genera) metagenomes from 15 and 16 publicly available datasets, respectively (Extended Data Fig. 1a and Supplementary Table 2), showing number of studies (c) and number of samples (d). e, From this metagenomic dataset, we reconstructed 13,446 MAGs, of which 1,524 were from Tara Pacific metagenomes (mostly from fire corals). In addition, we collected 103 isolate genomes from 26 studies (Supplementary Table 4). f, All genomic data were compiled to generate the RMD at https://rmd.microbiomics.io. Map in a adapted from Esri (https://server.arcgisonline.com/arcgis/rest/services/World_Topo_Map/MapServer). Imagery in this work is owned by Esri and its data contributors and are used herein with permission. Copyright © 2026 Esri and its data contributors. All rights reserved.
Applying a previously benchmarked bioinformatic workflow24 to this dataset, we reconstructed 13,446 coral- and sponge-associated MAGs from bacteria and archaea (Fig. 1e and Extended Data Fig. 1b). Of the 2,046 coral-associated MAGs, 1,964 were from reef-building corals and 1,524 originated from the Tara Pacific expedition28. With 57% (1,171) of all coral-associated MAGs, fire corals contributed more microbial genomes than stony (39%; 793) and soft (4%; 72) corals combined (Supplementary Table 3). Focusing on the Tara Pacific samples (equal sampling effort across coral hosts), we reconstructed 1,170 genomes from Millepora samples, 305 from Porites samples and 48 from Pocillopora samples. Although these numbers do not necessarily reflect microbial species richness, they are congruent with the observed differences across these hosts based on 16S rRNA gene sequencing results9. Furthermore, linking these genomic data to transmission electron microscopy (TEM) images, we found the high number of MAGs from fire corals to correspond to a high load of extracellular microorganisms (Fig. 1b and Extended Data Fig. 1c).
Overall, our efforts increased the number of available coral-associated MAGs tenfold (Supplementary Table 3). The data analysed in this study, which also includes 103 available isolate genomes (Supplementary Table 4), are referred to as the Reef Microbiomics Database (RMD) (Fig. 1f; Methods; ‘Data availability’).
Genomic novelty and host-specificity
Establishing the RMD enabled us to capture systematically the number of microbial species we had reconstructed genomes for and to assess the degree to which these species lacked previous genomic information. Specifically, we annotated all genomes using the Genome Taxonomy Database (GTDB; r207) toolkit and clustered genomes using a 95% whole-genome average nucleotide-identity threshold29, which defined 4,224 species-level clusters (species). Close to 90% (3,774) of all species, and 99% (638) of the 645 identified in Tara Pacific corals, were not present in the GTDB (Fig. 2a and Supplementary Table 3). Three-quarters of the species represented in the RMD had their genomes reconstructed from sponge metagenomes (3,206 versus 971 reconstructed from coral metagenomes) and less than 1% (36) of all microbial species were shared between sponge and coral metagenomes (Fig. 2a and Supplementary Table 3). Within corals, we reconstructed genomes from 516, 460 and 26 microbial species from stony, fire and soft corals, respectively (Fig. 2b). Stony and fire corals shared as few as 37 microbial species, and no species were shared with soft corals. More specifically, 95% of microbial species were unique to a particular host genus (Supplementary Table 3).
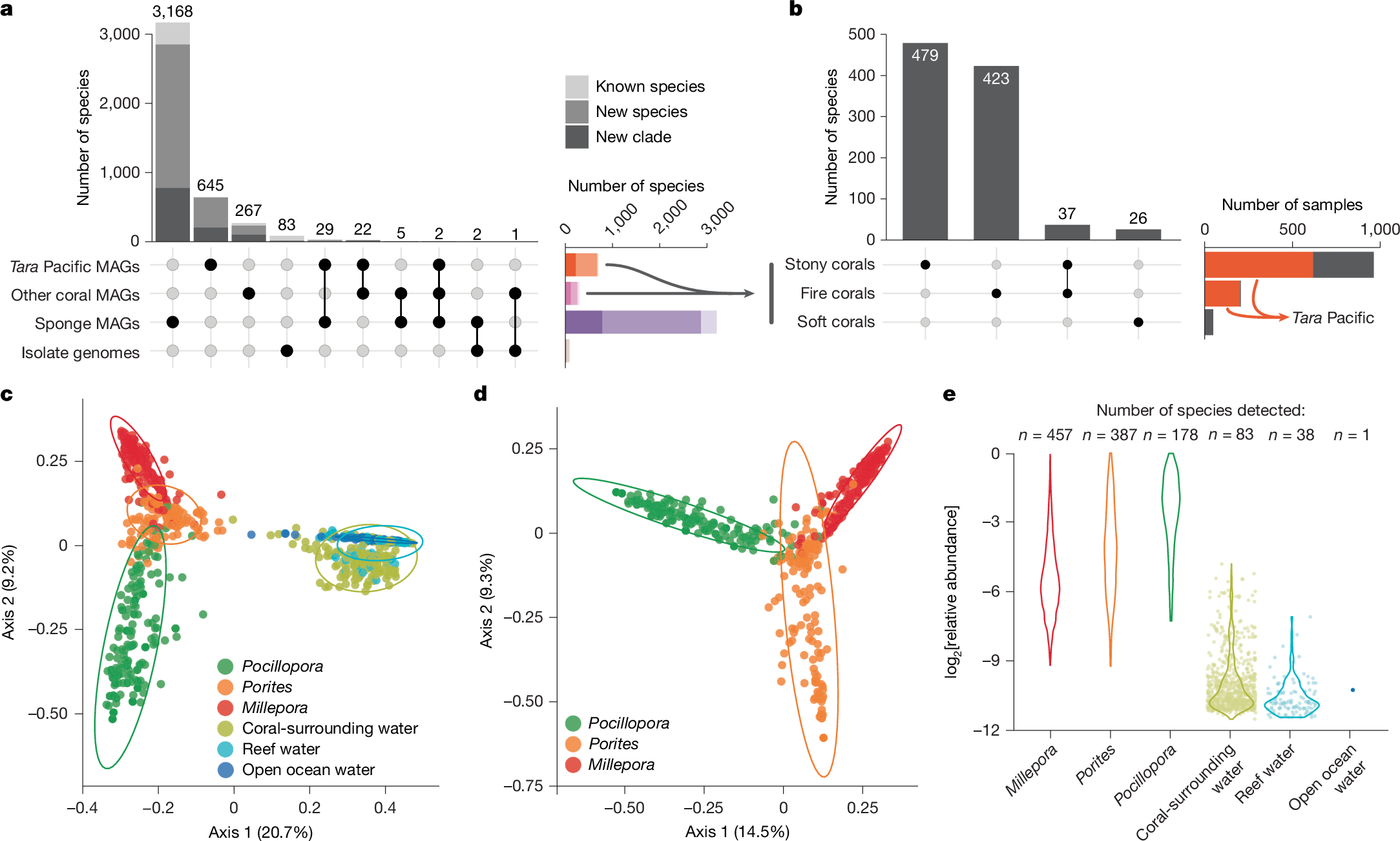
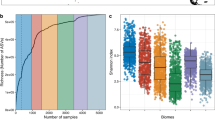
a, By grouping the 13,549 genomes in the RMD at the species-level (95% average nucleotide identity), we found them to represent 4,224 microbial species. Taxonomic annotation with the GTDB (r207) showed that close to 90% (3,774) of all species, and 99% (638) of those reconstructed from Tara Pacific corals, represented new species or clades at higher taxonomic ranks. b, The species displayed high specificity for the different types of coral (stony, fire and soft). c, A Jaccard distance-based PCoA of microbial species detected across the Tara Pacific coral and seawater metagenomes showed a clear separation between the microbiomes of corals (Pocillopora n = 207, Porites n = 177, Millepora n = 201) and seawater (coral-surrounding water n = 181, reef water n = 95, open ocean water n = 111) (PERMANOVA, P value ≤ 0.001, R2 = 0.31). d, Likewise, the coral metagenomes from the three coral genera (Pocillopora, Porites, Millepora) targeted during the Tara Pacific expedition differed significantly (PERMANOVA, P value ≤ 0.001, R2 = 0.18). e, The number and abundance of coral-associated microbial species decreased in seawater samples as a function of distance to the coral host. Individual data points are shown for groups containing fewer than 100 data points.
Beyond reconstructing genomes, we sought to validate this pervasive host-specificity by comparative compositional profiling of the host-associated microbial species identified here. To this end, we used a single-copy-marker-gene-based method30 to determine the species-level taxonomic composition of all (1,603) host-associated metagenomes, 387 seawater metagenomes collected during the Tara Pacific expedition28 (Fig. 1b) and 84 seawater metagenomes from previous coral- and sponge-focused studies that we included in our analysis (Methods). Overall, the microbial species profiles of coral metagenomes were not only distinct from those of seawater metagenomes (Fig. 2c and Extended Data Fig. 1d,e) but were also host specific (Fig. 2d).
Finally, we used the Tara Pacific dataset to test the detectability of coral-associated microorganisms in seawater sampled at increasing distances from their host, that is, coral-surrounding water, reef water and open ocean water. We detected as little as 20% of all microbial species from the coral metagenomes in the water samples. Furthermore, both the number and relative abundance of the detected species decreased with distance from the sampled coral colony within the reef, and they were barely detected in the open ocean (Fig. 2e).
Together, our results complement previous metagenomic efforts to reconstruct microbial genomes from diverse environments22