Substrate selectivity of the human RNA m5C methyltransferase NSUN2
Abstract
Specific deposition of RNA modifications is important for regulating gene expression1,2. 5-Methylcytosine (m5C) is a common epitranscriptomic modification, and NSUN2 is a key enzyme responsible for m5C methylation of various types of RNA. Dysregulation of NSUN2 is associated with numerous diseases, including cancers and neurological disorders3. The versatility of NSUN2 complicates our understanding of its substrate specificity and molecular roles in biology and disease. Here we show how NSUN2 interacts with RNA substrates at distinct stages of its catalytic cycle to modify cytidines. Furthermore, we show the role of RNA structure in facilitating NSUN2 activity at multiple tRNA positions. We identify RNA duplexes surrounding the m5C modification site as crucial recognition elements for methylation, which enabled us to derive a minimized substrate that captures the preferred features of an NSUN2 substrate—a dual-stem structure containing the CNNRR motif at the 5′ end of the first stem. Insights into the mechanisms underlying substrate-specific NSUN2 enzymatic activity provide opportunities for understanding and therapeutically targeting NSUN2-dependent methylation. Overall, our work highlights the roles of RNA structure and sequence in defining substrate specificity and regulating RNA-modifying enzymes.
Similar content being viewed by others
NSUNs-driven dysregulation: the next frontier in targeted cancer therapy?
NSUN2-mediated RNA m5C modification drives multiple myeloma progression by enhancing the stability of HIP1 mRNA

A cohort of mRNAs undergo high-stoichiometry NSUN6-mediated site-specific m5C modification
Main
RNAs are decorated with various chemical modifications that determine their fate, and specific m5C deposition is important for regulating gene expression necessary for cell proliferation and differentiation, development, stress response and immunity3,4. The NOP2/Sun (NSUN) family of methyltransferases add m5C modifications to RNAs5, which can then be detected by reader proteins to regulate RNA function, transport or stability and also be removed by TET enzymes or ALKBH1 (refs. 6,7,8,9). Thus, m5C is a dynamic epitranscriptomic modification that provides a handle for regulating RNAs post-transcriptionally.
NSUN2 is the primary m5C writer for mRNAs, and its dysregulation has been implicated in numerous human diseases10,11,12,13. Loss-of-function mutations in NSUN2 cause intellectual disability and Dubowitz-like syndrome, as NSUN2 is important for neural circuit formation and synaptic function14,15,16,17. NSUN2 also regulates ageing, tissue regeneration and tumourigenesis18,19,20,21,22,23,24. Notably, NSUN2 copy number is amplified in more than 10% of non-small-cell lung cancers25 (Extended Data Fig. 1a), and dysregulated m5C metabolism also promotes leukaemogenesis26. Therefore, NSUN2 is an important biological regulator and a potential therapeutic target.
Each m5C writer exhibits distinct substrate preferences5. NSUN2 is an especially versatile methyltransferase that can modify many types of RNA, including mRNAs, tRNAs, various ncRNAs and viral RNAs27,28,29. Even within tRNAs, NSUN2 can methylate multiple positions30,31,32. However, the mechanism underlying the specific and regulated methylation activity of NSUN2 remains unknown, and transcriptome-wide studies yield no clear consensus motif10,11,13,33. Deciphering the molecular mechanisms underlying NSUN2 activity is important for understanding its broader impact on human health and pathology.
Here, we describe the mechanism by which NSUN2 specifically methylates RNA. We present a high-resolution cryogenic electron microscopy (cryo-EM) structure (2.57 Å) of an NSUN2–RNA reaction intermediate. Moreover, we provide eight more cryo-EM structures of NSUN2–substrate complexes at various stages of the catalytic cycle to explain how an interplay among different RNA conformational states, cofactor binding and specific protein–RNA contacts drives reaction progress. Furthermore, we show how the methyltransferase can modify multiple sites within the same tRNA scaffold while maintaining specificity. We identify a dual-stem structure with a bulge-like linker and an m5CNNRR motif as the preferred features of NSUN2 substrates. We introduce a simplified, minimal substrate to demonstrate how NSUN2 targets are more generally recognized. Overall, our findings show the molecular basis of m5C modification, providing a framework for understanding its role in biology and disease.
Cryo-EM structure of NSUN2–tRNA
To understand the mechanism of NSUN2-mediated m5C modification of RNAs, we aimed to obtain an atomic model of the enzyme–substrate complex. We selected tRNAs as they are well-established substrates of NSUN2 (refs. 30,31) and are known for their stable, conserved fold. We purified recombinantly expressed wild-type full-length human NSUN2 protein and reconstituted its methylation activity with purified, in vitro transcribed tRNAs (Fig. 1a and Extended Data Fig. 1b). tRNALysCTT contains a known NSUN2 methylation site at Cyt48 in the variable region, and is robustly methylated in vitro. By contrast, tRNAIleTAT is a poor methylation substrate because it lacks a cytosine in this region. When Cyt48 of tRNALysCTT is mutated to uridine, the catalytic activity becomes undetectable, although the binding affinity remains comparable to that of wild-type substrates in the electrophoretic mobility shift assay (EMSA; Extended Data Fig. 1c,d), indicating that stable protein–RNA complex association is insufficient to drive methylation without cytosine at position 48 in tRNALysCTT. Thus, we successfully reconstituted the specific methylation activity of NSUN2 with a tRNA substrate.
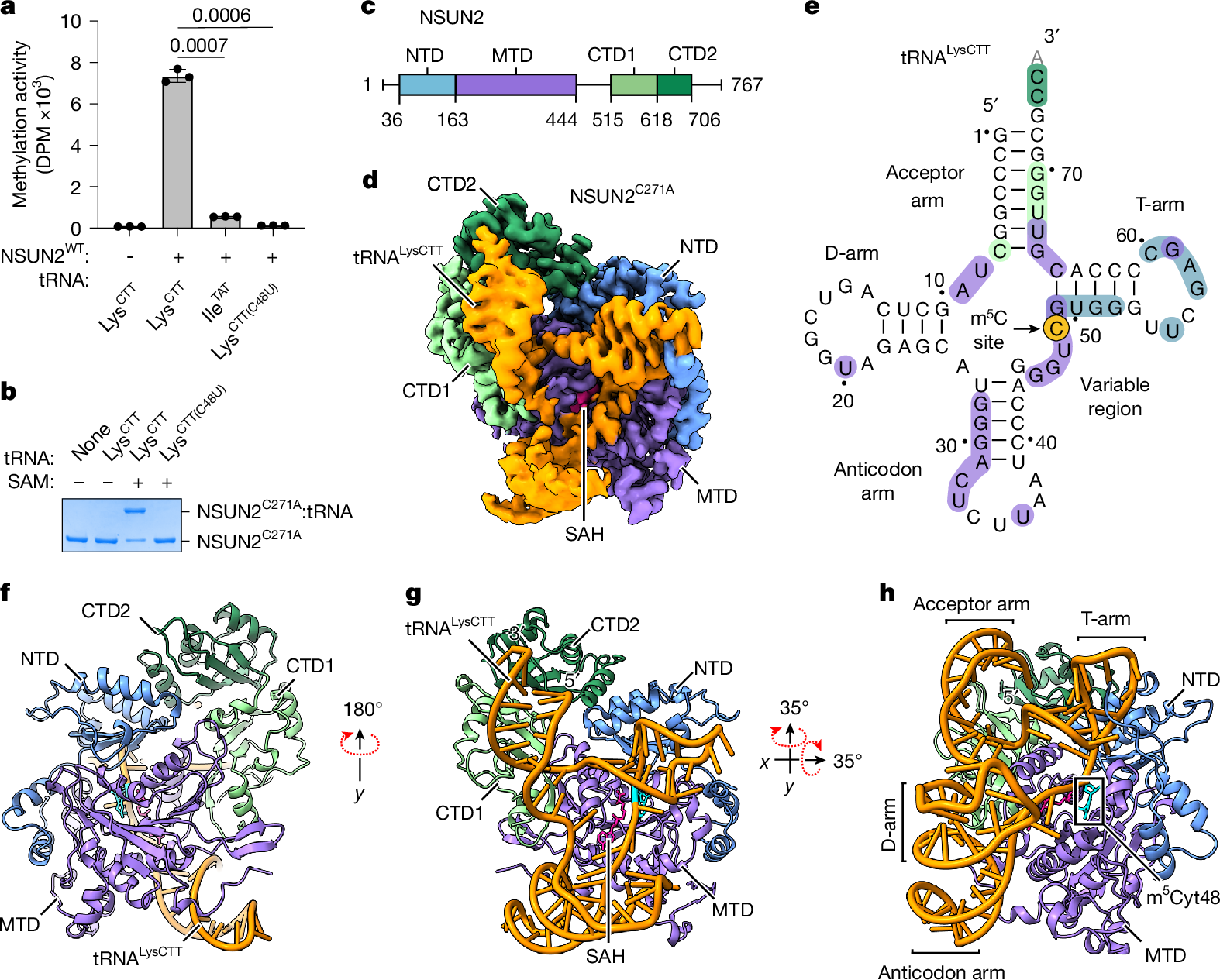
a, In vitro methylation activity of full-length wild-type (WT) human NSUN2 with indicated tRNA substrates, shown as mean ± standard deviation (s.d.) (n = 3). Statistical significance is indicated (two-tailed unpaired t-test). DPM, disintegrations per minute. b, In vitro crosslinking activity of NSUN2C271A with indicated tRNA substrates. Representative SDS–PAGE gel from three replicates is shown. For gel source data, see Supplementary Fig. 1. c, Domain organization of human NSUN2. d, Sharpened cryo-EM density map of the NSUN2C271A–tRNALysCTT complex with bound SAH (pink) and RNA (orange) in the D-arm conformation. The NSUN2 portion is coloured by domain as marked in c. e, Cloverleaf diagram of tRNALysCTT used in cryo-EM. Protein–RNA contacts (distance <3.8 Å) are coloured by the interacting protein domain, as in c, with the target cytosine highlighted in yellow. Unresolved nucleotide 76 is shown in grey. f–h, Models of the NSUN2C271A–tRNALysCTT–SAH complex in the D-arm conformation in cartoon representation from three different views, with SAH as pink sticks and m5Cyt48 as cyan sticks.
To gain insight into the catalytic activity of NSUN2, we sought to stabilize the reaction intermediate by leveraging its unique chemical mechanism. According to the proposed mechanism conserved for m5C methyltransferases34,35, NSUN2 forms a covalent linkage with the substrate cytosine base through a cysteine (C321), activating the methyl transfer from S-adenosylmethionine (SAM) to the cytosine and yielding S-adenosylhomocysteine (SAH) as a by-product; subsequently, a second cysteine (C271) is necessary to resolve the catalytic intermediate (Extended Data Fig. 1e,f). We mutated C271 to alanine (C271A) to inhibit the dissolution of the protein–RNA conjugate, as an intermolecular crosslink would increase the stability of the protein–RNA complex, enabling us to determine a higher-resolution complex structure and reveal a view of the catalytic intermediate. NSUN2–tRNA conjugate formation requires SAM and Cyt48, and the specific crosslinking activity is readily detectable as the conjugate is resistant to denaturing conditions (Fig. 1b).
We used single-particle cryo-EM to determine the structure of an NSUN2C271A–tRNA crosslinked complex at 2.57 Å resolution (Fig. 1c–e, Extended Data Table 1 and Extended Data Figs. 1 and 2). The overall architecture of the complex shows that NSUN2 contains four domains: an N-terminal domain (NTD), a methyltransferase domain (MTD) and two C-terminal domains (CTD1 and CTD2). The catalytic MTD is the largest domain and organizes the protein by making direct contact with NTD and CTD1, and CTD2 is wedged between CTD1 and NTD (Fig. 1f). NSUN2 uses all the domains to make extensive contact (2,878 Å2) with the tRNA, and most of the tRNA interacts with NSUN2, except for the D-arm (Fig. 1g,h). The lack of substantial protein contact probably leads to conformational variability in the D-arm; we observe two main classes of particles: one with a visible D-arm and the other with a disordered D-arm (Extended Data Fig. 1g,h). We also observe that the interactions between NSUN2 and the anticodon arm are distinct in these two conformations (Extended Data Fig. 1i,j), suggesting that the anticodon arm interacts with the protein more weakly. Nevertheless, despite weaker contact with D- and anticodon arms, the tRNA molecule is stably bound to NSUN2 to produce a high-resolution map of the RNA–protein complex (Extended Data Fig. 2).
A striking feature of the NSUN2–tRNA conjugate structure is the unusual conformation of the bound tRNA. In the NSUN2-bound state, the tRNA no longer adopts its canonical L-shape that depends on the contact between the T- and D-loops to form the elbow (Fig. 2a and Extended Data Fig. 1k). Structural superimposition indicates a large amount of unfolding, in which Gua19N1 and Cyt56N3, which typically form a hydrogen bond as part of the kissing loop interaction between the D- and T-loops, become separated by about 35 Å. This conformational change of the tRNA is probably stabilized by extensive protein–RNA contacts. Although NSUN2 envelops most of the tRNA by a large basic surface (Fig. 2b), the most intimate contact is observed near the NSUN2 regions of higher evolutionary conservation that bind the T-arm and parts of the acceptor arm (Fig. 2c–e). The T-stem is recognized by a row of basic side chains from the NTD that recognizes the pattern of phosphates from the duplexed RNA, ensuring a stem structure adjacent to the methylated cytidine (Fig. 2f and Extended Data Fig. 1l). Here, the guanidinium groups of R133 and R137 also contact N7 of guanine bases, which would cause purines to be preferred at these positions. The unpaired nucleotides in the T-loop form several favourable contacts with NSUN2, through ring-stacking and hydrophobic interactions, along with a few electrostatic contacts (Fig. 2g and Extended Data Fig. 1m). The base of the acceptor arm is buttressed by the MTD helix (α3) that provides a platform for the RNA duplex, and the phenol group of Y222 stabilizes Gua66 by ring stacking (Fig. 2h and Extended Data Fig. 1n). The backbone edge of the duplex RNA in the acceptor arm makes favourable contacts with the CTD1 (Fig. 2i and Extended Data Fig. 1o). At the 3′ end of the acceptor arm, the single-stranded RNA (ssRNA) is ordered enough to be modelled, although less defined than most of the RNA, suggesting that it may have loose interactions with the CTDs (Extended Data Fig. 1p). The key RNA–protein contacts can be summarized using a schematic (Fig. 2j). Several point mutations in these contact areas can diminish productive methylation, in agreement with our structural observations (Fig. 2k). The R133Q mutation has been observed in tumour samples36 (Fig. 2f), and reduced methylation activity may contribute to certain cancers. Therefore, the structure-guided biochemical analysis suggests that the NSUN2–tRNA complex is primarily stabilized by RNA structure-dependent interactions, including electrostatic interactions with the phosphate edges of the duplex (acceptor arm and T-arm) and sequence-independent stacking interactions with the T-loop.
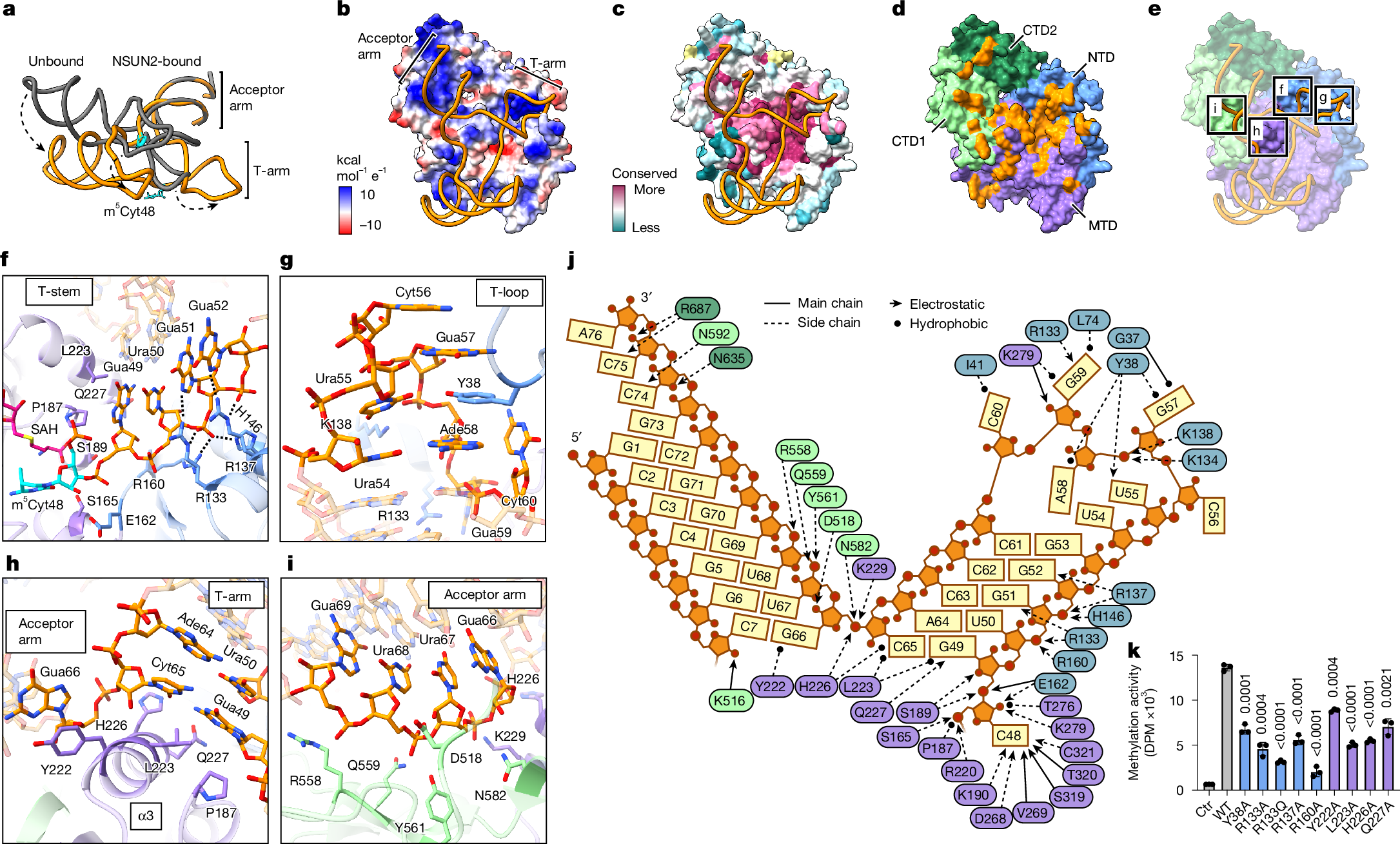
a, Alignment of unbound (PDB: 1FIR, grey) and NSUN2-bound (PDB: 9Z2N from this study, orange) tRNALysTTT by their acceptor arms. Dashed arrows show the conformational differences in key structural elements. b,c, Models of the NSUN2C271A–tRNALysCTT complex with RNA shown as ribbon and protein in surface representation, coloured by vacuum electrostatic potential (b) or evolutionary conservation score (c). Conservation score per residue calculated using ConSurf, with yellow indicating insufficient data. d, NSUN2 surface representation coloured by domain, except for the residues within 3.8 Å of RNA that are shown in orange. e, Similar to d, protein coloured by domain, with RNA–protein interfaces highlighted with black boxes and panel labels. f–i, Close-up views of protein–RNA interfaces highlighted in e. Protein side chains within 3.8 Å of RNA are shown as sticks, and electrostatic contacts (<3.8 Å) are indicated by black dashed lines. Cyt48 is shown in cyan. j, Schematic of protein–RNA contacts (<3.8 Å) in T- and acceptor arms with protein residues coloured according to d. Arrows indicate electrostatic (triangle) or hydrophobic (circle) interactions between main chain (solid) or side chain (dashed) protein atoms with RNA. k, In vitro methylation activity of NSUN2 mutants with substitutions in NTD (blue) or MTD (purple), with wild-type tRNALysCTT, shown as mean ± s.d. (n = 3). Ctr, no enzyme control. Significance tests are comparisons with NSUN2WT (two-tailed unpaired t-test).
Conformational changes during catalysis
Our NSUN2–tRNA conjugate structures provide a high-resolution view of the NSUN2 catalytic pocket in action, revealing the reaction intermediate (Fig. 3a and Extended Data Fig. 1q). The methylated cytosine ring and the by-product SAH are deeply buried in the catalytic pocket. Our maps are sufficiently resolved to model the SAH and the protein–RNA crosslink between Cyt48 and the catalytic residue C321. An intricate network of polar contacts and van der Waals interactions determines the conformations of the cofactor and the methylated cytosine ring. The specificity for the cytosine base is conferred through recognition of N4 by main-chain carbonyl interactions with V269 and S319 (Extended Data Fig. 1r). RNA drapes over the bound SAH, suggesting the bound cofactor helps organize the RNA near the catalytic pocket (Extended Data Fig. 1s). As expected for residues responsible for precise chemistry, point mutations of the amino acids involved in coordinating the catalytic pocket are deleterious for methylation activity, including t