Human haematopoietic stem cells remember inflammatory stress
Abstract
Inflammation activates blood cells, contributing to ageing and malignancy1,2,3. Haematopoietic stem cells (HSCs) survive a lifetime of infection to sustain life-long haematopoiesis1,2,3,4,5,6,7,8,9, but how human HSCs respond and adapt to inflammatory stress is largely unknown. Here, to empirically understand this adaptation, we developed xenograft inflammation–recovery models and performed single-cell multiomics on xenografted human HSCs. Two transcriptionally and epigenetically distinct HSC subsets were identified with one, termed HSC inflammatory memory (HSC-iM), retaining a molecular memory of previous inflammatory treatments. The HSC-iM subset exhibited quiescence and restrained haematopoietic output. Molecularly, the HSC-iM program was enriched in HSCs from adult and paediatric samples across conditions ranging from COVID-19 recovery, sickle cell disease, ageing and clonal haematopoiesis, establishing both the validity of our xenograft models and the physiological relevance of HSC-iM. Clonal haematopoiesis mutations in HSC-iM attenuated the effects of inflammatory stress by promoting HSC activation and differentiation. Moreover, transmission of the pro-inflammatory HSC-iM transcriptional program to differentiated immune progeny was demonstrated in xenograft and physiological settings. Finally, HSC-iM program enrichment in circulating blood cells was associated with a heightened risk score for all-cause mortality in population cohort analyses, underscoring the clinical relevance of this newly identified HSC subset in characterizing heterogeneous health outcomes across a lifetime.
Similar content being viewed by others
Inflammageing and clonal haematopoiesis interplay and their impact on human disease
Systemic and local regulation of hematopoietic homeostasis in health and disease

Chemically defined cytokine-free expansion of human haematopoietic stem cells
Main
Humans have enormous demand for haematopoietic output that is met by a complex cellular hierarchy with HSCs at its apex10. Mature blood cells with a finite lifespan are continuously replenished by bone marrow haematopoietic stem and progenitor cells (HSPCs) producing 3 million cells per second in human adults, via a tightly controlled process11. Human HSCs are both genetically and functionally diverse with 50,000–200,000 HSCs contributing to haematopoiesis at any one time12. With age, HSCs show functional decline1,9, have a marked decrease in clonal diversity7, a consequent increase in incidence of clonal haematopoiesis (CH), and an increased risk for blood malignancy13. How daily blood production is coordinated with HSC pool maintenance over a lifespan is unclear, especially when exposed to inflammatory stressors, for example, recurrent infections, which can induce HSC activation3,4,14,15. In addition, HSC fate transitions from quiescence towards activation are essential for HSC function, but can be dysregulated with ageing and inflammatory stress16,17,18,19,20. Finally, HSCs are not homogeneous; HSCs exhibit transcriptional, epigenetic and functional heterogeneity, where inflammatory pathways are a major source of variation14,18,19,20,21,22. Together, this indicates that heterogeneous cellular responses to inflammation within the human HSC pool might drive ageing phenotypes in response to life-long inflammatory insults9,23,24.
Human lineage tracking shows that pre-leukaemic CH mutations arise decades before disease detection25,26,27,28. CH mutation-bearing clones occur ubiquitously in adults older than 60 years of age, although only a fraction expand to reach detectable clone size13. Crucially, mechanisms regulating CH clone size are poorly understood. Mouse models have shown that inflammation activates HSCs, consequently impairing differentiation and self-renewal1,2,3,4,5,15,29,30, affecting clonal selection in CH6,29,31. Some inflammatory responses include activation of target genes downstream of tumour necrosis factor (TNF) via NF-κB, known to regulate HSC survival in both mouse and human settings3,14. Repeated inflammatory challenges have also been associated with sustained epigenetic changes30 and accelerated ageing of mouse HSCs1,4, as well as expansion of clones bearing CH-associated mutations5,6,29,31. However, given the heterogeneity of the HSC compartment, it is unclear whether all HSCs, especially in humans, respond equally to inflammation. If HSCs respond heterogeneously, it is also unclear whether this would affect clonal selection such as in CH.
Inflammatory response priming in LT-HSC
Human HSC can be deeply quiescent or cell-cycle primed; states tied to variations in regeneration kinetics, 3D chromatin architecture and endolysosomal activity14,19,20,21. We performed single-cell transcriptional profiling of cord blood (CB) Lin−CD34+CD38−CD45RA−CD90+CD49f+ long-term HSCs (LT-HSCs) and used non-negative matrix factorization (NMF) to explore transcriptional heterogeneity among 3,381 LT-HSCs at homeostasis (Fig. 1a). Programs of quiescence (Fig. 1b,c) and inflammatory signalling (Fig. 1d,e) were identified (Supplementary Tables 1 and 2). These were independently validated in single-nucleus gene expression and chromatin accessibility profiles (hereafter, scMultiome) of 15,590 CB HSCs and multipotent progenitors (HSC–MPPs) as identified using BoneMarrowMap32 (Extended Data Fig. 1a–h and Supplementary Table 3). The inflammatory program was associated with chromatin accessibility of AP-1 and NF-κB transcription factor-binding motifs (Extended Data Fig. 1i–k). To jointly evaluate the relevance of the distinct inflammation and quiescence programs identified by NMF within these datasets, ‘meta-programs’ were defined by consensus; these also showed stronger enrichment for inflammation-specific and quiescence-specific programs, respectively (Supplementary Table 4 and Supplementary Note 1). These data corroborate previous findings19,20,21,22 of variable levels of transcriptional inflammatory priming within individual CB HSC-MPPs.
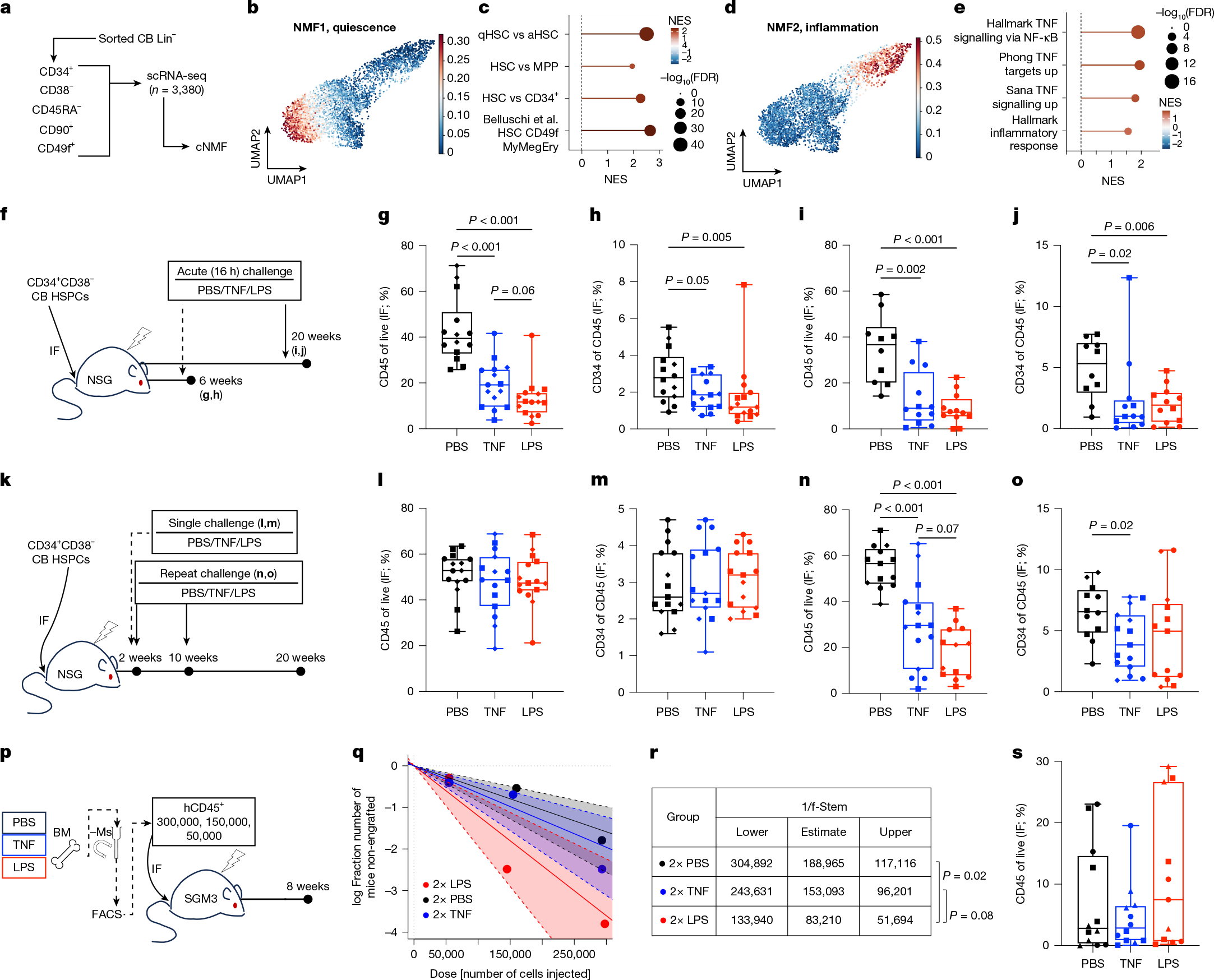
a, CB LT-HSCs were subjected to scRNA-seq. Consensus NMF (cNMF) was performed on 3,381 cells. b–e, Projection of a cNMF transcriptional program of quiescence (NMF1; b) or inflammation (NMF2; d), as defined by GSEA enrichment (c,e, respectively). aHSC, activated HSC; FDR, false discovery rate; MyMegEry, myeloid-megakaryocyte-erythroid lineage potential; NES, normalized enrichment score; qHSC, quiescent HSC; UMAP, uniform manifold approximation and projection. f–j, NSG mice xenografted with CB-derived CD34+CD38− cells were challenged with TNF or LPS at either 6 weeks (g,h; n = 14 for the PBS and n = 15 for the TNF and LPS groups across 3 independent pools) or 20 weeks (i,j; n = 10 for the PBS and n = 12 for the TNF and LPS groups across 2 independent pools) post-transplant. Total human engraftment (g,i) and progenitors (h,j) 16 h after inflammatory challenge or vehicle were measured. IF, injected femur. k, Schematic of the inflammation–recovery xenograft models. l,m, Human engraftment (l) and progenitor composition (m) 18 weeks after a single challenge (n = 15 mice per group across 3 independent CB pools). n,o, Human engraftment (n) and progenitor composition (o) 10 weeks after repeated challenge (n = 13 mice in PBS and LPS groups, and n = 15 in the TNF group across 3 independent CB pools). p, Schematic of the secondary transplantation with limiting dilution of human leukocytes from repeat-challenged xenografts. BM, bone marrow; FACS, fluorescence-activated cell sorting; −Ms, mouse depletion. q,r, Stem cell frequency (f-Stem) estimates for p. Dashed lines and shaded regions show 95% confidence intervals for each group. s, Human engraftment from p transplanted with 300,000 hCD45+ cells (n = 12 for PBS and TNF groups and n = 11 for the LPS group across 3 independent pools). Engraftment data are presented as boxplots: quartiles indicate a line at the median; the whiskers show the range; individual points are per mouse; and the symbols reflect independent CB pools. Engraftment data were compared with pairwise two-sided Mann–Whitney tests, whereas chi-squared tests were used for stem cell frequencies; P < 0.1 is shown numerically, whereas P > 0.1 is not shown.
Modelling human HSC inflammation in vivo
To investigate how CB HSCs would functionally respond to inflammatory stress, we developed a xenotransplantation model of inflammation and focused on the impact of lipopolysaccharide (LPS), which mimics sepsis33, and TNF, which has been associated with deleterious effects on human health in the context of advanced age, COVID-19 and sepsis34,35,