Darkness and body size shaped end-Cretaceous marine extinction patterns
Abstract
The Chicxulub asteroid impact at the Cretaceous–Paleogene (K–Pg) boundary (66 Ma) is thought to have caused the extinction of around 75% of species in the fossil record by triggering catastrophic environmental changes1. However, despite decades of research, the mechanisms linking the environmental changes to the selective extinction patterns observed in the marine fossil record remain unresolved. Here we use a global trait-based ecosystem model2,3 to establish this causality for the marine plankton community beyond the fossilized groups. Our model simulates diversity dynamics during the initial 100 years after the K–Pg boundary and represents explicitly extinction based on biomass thresholds that scales with body size. Under K–Pg climatic forcings, the model reproduces successfully key observed extinction patterns, including the high vulnerability of planktic foraminifera and other zooplankton, the survival of small mixotrophs4 and phytoplankton5,6, and potential for reduced diversity loss in high-latitude settings7. Our analysis suggests that impact-driven darkness and body-size-dependent extinction thresholds drove most of the observed extinction patterns. These results suggest that plankton ecologies enhance survival through differences in energy demand and acquisition. Our study bridges the gap between fossil evidence of extinction patterns and the K–Pg impact winter hypothesis, highlighting the value of trait-based models for understanding past biodiversity crises.
Similar content being viewed by others
Seasonal calibration of the end-cretaceous Chicxulub impact event

Major restructuring of marine plankton assemblages under global warming
Regional restructuring in planktic foraminifera communities through Pliocene-early Pleistocene climate variability
Main
The Cretaceous–Paleogene (K–Pg) boundary (66 Ma) is marked by a mass extinction event1 that altered Earth’s terrestrial and marine biosphere profoundly. Both the emplacement of the Deccan Trap8 and the Chicxulub asteroid impact on the Yucatán carbonate platform were recognized as the potential drivers of this mass extinction. However, overwhelming evidence supports the latter triggering the marine extinction through abrupt environmental changes9,10. The bolide impact and associated wildfires released silicate dust, soot and sulfur aerosols into the atmosphere9, blocking solar radiation and causing the reduction of light and global cooling11. Simultaneously, global wildfires and the vaporization of carbonate-rich target rock increased CO2 concentrations by approximately 700 ppm across K–Pg12.
Despite advances in reconstructing the sequence of events across the K–Pg boundary, it is still unclear how environmental changes caused observed extinction patterns13. In the ocean, nearly all nannoplankton, planktic foraminifera, all rudist bivalve and ammonoid cephalopod molluscs went extinct14,15,16. By contrast, dinoflagellates, diatoms, radiolarians and benthic foraminifera were less affected17,18. Notably, high-latitude nannoplankton7, particularly in the Southern Ocean, have exhibited lower extinction rates than their low-latitude counterparts, and a similar pattern might exist for planktic foraminifera19. However, Northern Hemisphere high-latitude data remain limited7 and such a latitude-dependent extinction was not found in molluscs20. Furthermore, surviving nannoplankton and foraminifera were small and opportunistic4,21, with similar size reductions found in other marine organisms22.
Several hypotheses have been proposed to explain these ecological and geographical selective extinction patterns across the K–Pg boundary. Alvarez and colleagues23 suggested a dramatic loss of marine primary production due to reduced solar radiation, leading to a cascading trophic collapse. However, subsequent observations show that the reduction in productivity across the K–Pg was relatively modest24 and spatially heterogeneous25. The basin-dependent productivity change across the K–Pg also does not match the latitude-dependent extinction25. Instead, ocean acidification might have contributed to the higher extinction rate of calcifying organisms compared with silicifying organisms26. However, ocean acidification at the K–Pg was limited12 compared with other geological events27, similar to the Paleocene-Eocene Thermal Maximum where comparable acidification levels did not result in a global extinction of calcifiers28. These studies show that a mechanistic understanding is still lacking to reconcile the ecological and geographical selectivity observed in the fossil record of the K–Pg extinction.
Mechanistic ecosystem and biogeochemical models within Earth system models provide a powerful tool for linking K–Pg environmental changes to marine plankton dynamics. However, existing marine biogeochemical models do not simulate mass extinction explicitly, as these models focus typically on biogeochemical cycling and allow plankton populations to recover immediately under favourable conditions, even from extremely low biomass levels, which could obscure true extinction4,29. Furthermore, extinction thresholds for different marine groups remain poorly constrained30, hindering the reproduction and consequent investigation of observed patterns of extinction selectivity.
Body size is recognized widely as a master trait that shapes organism biology and strongly influences extinction thresholds of marine organisms in the Phanerozoic31. Larger organisms tend to be more vulnerable during mass extinctions due to their higher energy demands, lower population density and slower mass-specific metabolic rates32,33. This suggests that body size-dependent extinction thresholds could provide a mechanistic link between climate change, plankton ecology and extinction risk, yet these are not explored for the end-Cretaceous crisis.
Here we use a size-based mechanistic ecosystem model (EcoGENIE) to investigate the causes of observed extinction patterns in plankton communities across the K–Pg. EcoGENIE resolves size-dependent plankton ecophysiological processes explicitly within a three-dimensional ocean circulation and biogeochemistry framework (Supplementary Fig. 1). To overcome limitations of previous modelling approaches, we implement an extinction mechanism in EcoGENIE based on a size-dependent biomass threshold, defined as the biomass of a single individual of a given size. Larger plankton thus have higher biomass threshold and higher extinction vulnerability. We initialize the model with a diverse plankton community (32 phytoplankton, 32 generic zooplankton, 32 mixotrophs and 16 foraminifera functional types across size classes; Supplementary Table 1)34,35. This model allows environmental conditions to select which Late Cretaceous plankton functional types to go extinct in response to K–Pg environmental changes (Fig. 1). Such an approach enables us to examine extinction selectivity across the entire plankton community, including those without mineralized shells and hence absent from the fossil record. Furthermore, the explicit inclusion of planktic foraminifera within the zooplankton group offers an opportunity to validate model results directly against fossil data.
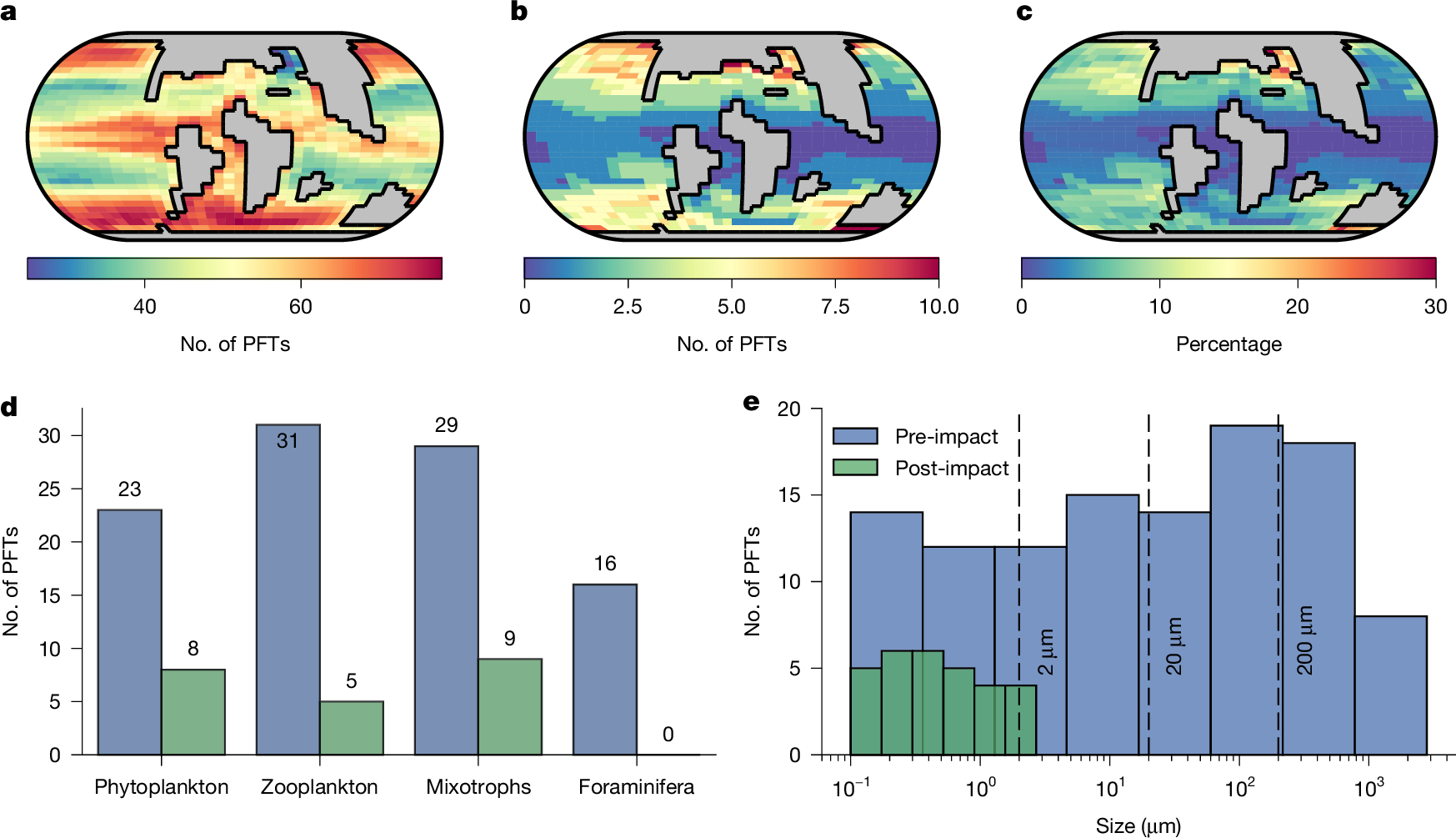
a,b, PFT richness before (a) and after (b) the K–Pg impact. c, Percentage of plankton survivors. d, Diversity of various PFTs before and after the K–Pg impact; this excludes the Arctic Ocean, which the model does not represent due to limited grid resolution. e, Size distribution across all existing plankton types before and after impact. Dashed vertical lines represent 2, 20 and 200 µm. Note that the colour scale varies between a, b and c to highlight geographical patterns.
Late Cretaceous climate and diversity
We applied the cGENIE model to a Late Cretaceous climate state (834 ppm CO2) and paleogeographic configuration (Supplementary Fig. 2 and Methods). The spin-up simulation produced a global mean sea surface temperature of around 26.4 °C, closely matching the latest data compilation based on TEX86 and δ18O proxies36 (Extended Data Fig. 1). The modelled deep-water temperature (deeper than 2,000 m) and surface pH are 9.3 °C and 7.7, respectively, both in line with Late Cretaceous oxygen and boron isotope estimates (9 °C and 7.7–7.8 pH units)12,37. In contrast to other models29,38, cGENIE succeeds in simulating a reduced equator-to-pole temperature gradient, in agreement with proxy data. Hence, it provides a realistic climatic environment to test extinction selectivity.
In the spin-up experiment, 99 plankton functional types survived, representing around 88% of the initial community. The emergent plankton diversity (defined as the coexisting number of plankton functional types) was highest in equatorial and coastal upwelling regions and at high latitudes (Fig. 1a). This spatial pattern aligns with the theory that the number of supported plankton size classes increases with nutrient supply rate39, which captures modern plankton observations successfully40. Therefore, although fossil data are spatially incomplete and biased towards groups with tests or shells, our model provides a plausible estimate of global plankton functional diversity in the Late Cretaceous.
Modelling the K–Pg extinction
We then perturbed the Late Cretaceous baseline simulation for 100 years following the impact, using a 7.30-h timestep for ecosystem dynamics. Three K–Pg climatic forcings derived from previous studies11,12 were applied to ensure consistency (Supplementary Information): (1) a CO2 pulse of 700 ppm, based on boron isotope data from foraminifers12; (2) a reduction in solar radiation (Extended Data Fig. 2); and (3) an ejecta-derived nutrient (iron and phosphorus) flux (Extended Data Fig. 3). The CO2 perturbation affects both climate and seawater carbonate chemistry (for example, pH); however, we note that potential acidification impacts on plankton are missing as the model does not incorporate directly the effect of pH on plankton physiology and biomineralization (Methods). The solar radiation forcing follows estimates from Senel and colleagues11, which combined sedimentological evidence with a climate model capturing the effects of sulfur, soot and silicate dust. The projectile-derived nutrient flux and distribution were re-gridded from a previous modelling study29, with peak deposition concentrated near the Chicxulub impact site.
The transient experiment shows that global annual mean sea surface temperature dropped from 26.4 °C to a minimum of 12.3 °C within the first 3 years post-impact (Fig. 2 and Extended Data Fig. 4). Recovery to pre-impact temperatures took around 30 years, resulting in a prolonged global impact winter, despite an intense CO2 release (1,750 Pg C; Fig. 2a). This abrupt cooling nearly eliminated the vertical seawater density gradient (Supplementary Fig. 3), leading to the global mean ocean mixed layer depth (MLD) deepening dramatically to 750 m after 2 years (Fig. 2c). Such a loss of a stratified ocean would have impacted the vertical niche separation of plankton dramatically. The deepest mixed layer occurs at approximately 60° N/S, similar to results from a more complex ocean model41 (Supplementary Fig. 4). This increased vertical mixing, combined with enhanced dust-derived nutrient fluxes, led to a substantial rise in surface nutrient availability (Fig. 2d), with modelled PO4 concentrations increasing by 17 times within the first 2 years following the impact (from 0.07 to 1.2 μmol P kg−1).
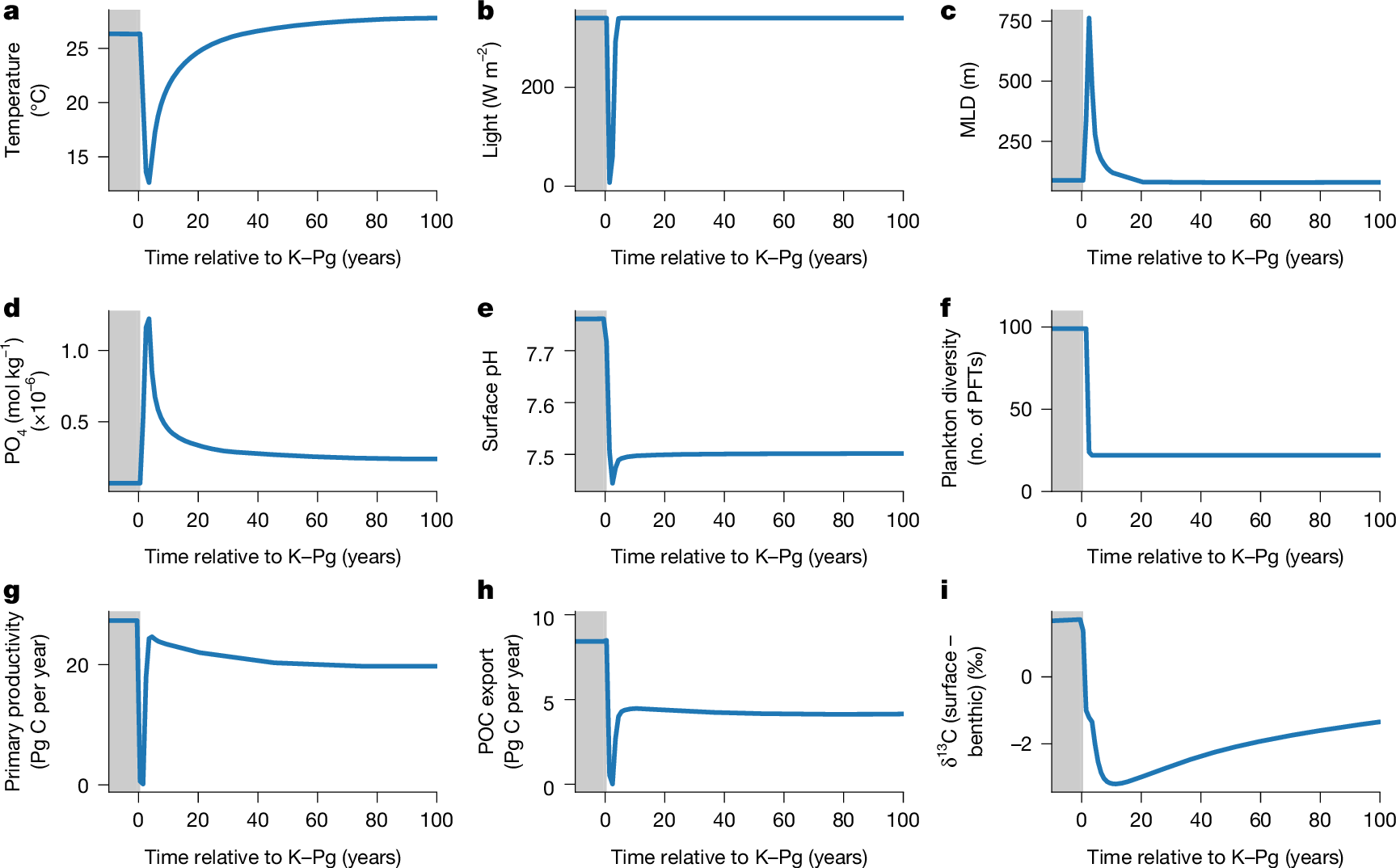
a, Global mean surface ocean temperature. b, Global mean insolation. c, Global mean MLD. d, Ice-free global mean phosphate concentration. e, Global mean ocean surface pH. f, Global coexisting PFTs. g, Total marine annual primary productivity. h, Annual particular carbon export (at 80.8 m). POC, particulate organic carbon. i, δ13C difference between surface (80.8 m) and benthic (deeper than 2,000 m) layers. Shaded areas, pre-im