Cellular water-potential sensing through biomolecular condensation
Abstract
Water molecules, as solvents for biomolecules, are essential to cells. The water potential of the cell decreases under water-deficient conditions1,2, yet how cells sense changes in water potential remains unknown. Here we identify a sterile alpha motif (SAM)-containing protein, SAM8, that undergoes water-potential-dependent condensation both in vivo and in vitro and is crucial for hyperosmotic stress tolerance and seed germination. We use biophysical techniques, in vitro reconstitution and bioimaging to demonstrate that SAM8 is strongly hydrated under normal water conditions, preventing its macroscopic condensation. A negatively charged patch determines SAM8 hydration by creating an electric field and micropolar environment. Water-deficient conditions weaken this hydration, thereby activating SAM8 condensation by reprogramming hydrogen bond, electrostatic and hydrophobic interactions. Furthermore, we demonstrate that SAM8 condensates selectively sequester RNA export factors, leading to nuclear retention of mRNAs and translational reprogramming under hyperosmotic stress. Our findings show a mechanism by which plant cells directly sense and respond to water status, shedding light on how they adapt to water deficit conditions.
Similar content being viewed by others
Macromolecular condensation buffers intracellular water potential
Probing the structure of water in individual living cells
Main
Water molecules are central to life, providing a solvent that maintains the functional structures and activities of biomolecules within cells. Cellular water molecules are bound by macromolecules, forming the hydration layer or freely diffuse in the bulk. These two portions of water are referred to as interfacial water and free water, respectively1. Water potential, which can be understood as the availability of free water, governs water uptake from the soil and transport within the plant3,4. The cellular water potential is sensitive to environmental fluctuations, particularly drought, high salinity and temperature stress1,2,5. Biomolecules in living cells are surrounded by at least a molecular layer of hydrated water6. Cellular water-potential reduction decreases hydration of biomolecules, compromising membrane integrity, disrupting the three-dimensional structures of proteins, impairing enzymatic activities, and so on7. Therefore, cellular water-potential sensing and response are crucial for plant development and adaptation to various environmental stresses, but the mechanisms underlying them remain unknown.
SAM8 undergoes water-dependent condensation
Biomolecular condensation is emerging as a key mechanism for sensing and responding to environmental stress8,9. Hyperosmotic stress often induces cell volume shrinkage, resulting in membrane tension changes, water-potential reduction and subsequent dehydration of biomolecules, increased molecular crowding and ion concentration10 (Fig. 1a and Extended Data Fig. 1a). Both nuclear and cytoplasmic proteins were reported to sense hyperosmotic stress by molecular crowding-dependent condensation in plants11,12. We reasoned that biomolecular condensation can be a way of sensing cellular water-potential changes because water tunes the strength of hydrophobic interactions, electrostatic interactions or the cation–π interactions, all of which are the driving force for condensation. Deuterium oxide (D2O) forms stronger hydrogen bonds than H2O, strengthening water–water interactions while reducing water–protein interactions13,14,15,16,17. Therefore, we used D2O treatment to reduce the water bound to proteins without affecting cell volume (Fig. 1a and Extended Data Fig. 1b).
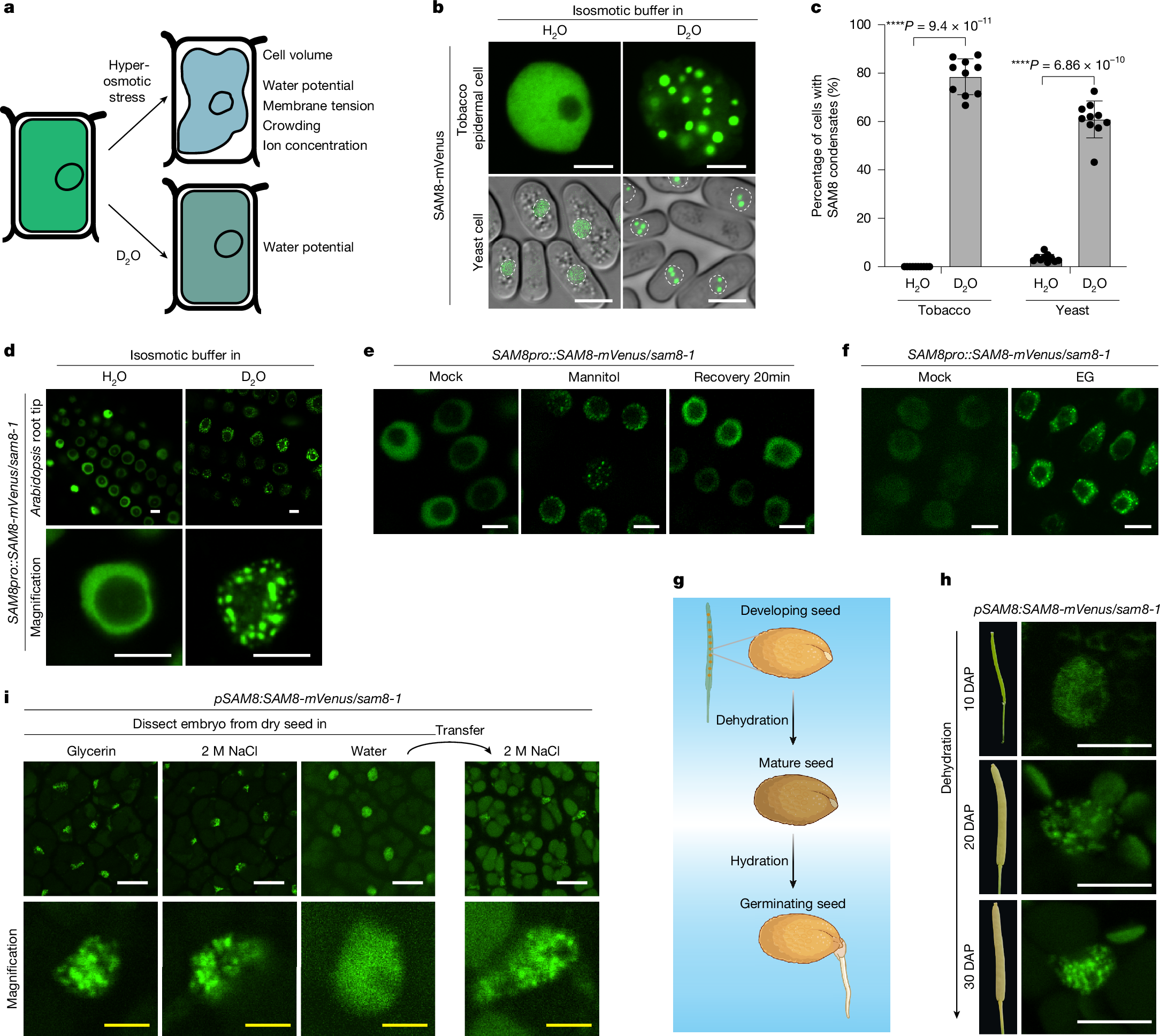
a, Illustration of the impact of osmotic stress and D2O on plant cells. b, Representative confocal images of tobacco epidermal cells (top) or Schizosaccharomyces pombe cells (bottom) expressing SAM8-GFP. The cells were treated for 10 min with an isosmotic buffer prepared with H2O or 90% D2O. The white dotted circles indicate the nuclei of yeast cells. Scale bar, 5 μm. c, Quantification of the percentage of cells containing SAM8 condensates as indicated. Error bars indicate mean ± s.d. (n = 10 independent experiments). d, Representative confocal images of Arabidopsis thaliana root tip cells that were treated with an isosmotic buffer prepared with H2O or 90% D2O. Scale bar, 5 μm. e,f, Representative confocal images of Arabidopsis root tip cells expressing SAM8-mVenus. The cells were treated with 0.3 M mannitol or 0.5 M EG for 10 min. Scale bar, 5 µm. g, Illustration of the dehydration and hydration process during seed development and germination. h, Representative confocal images of embryos dissected from the different stages of seeds expressing SAM8. DAP, days after pollination. Scale bar, 5 μm. i, Representative confocal images of embryos dissected from the dry and imbibed seeds expressing SAM8. Scale bar, 10 μm (white); 2 μm (yellow). Representative images of n independent experiments (n = 3 (d–f, h and i)). Asterisks indicate significant differences (two-tailed t-test).
To understand how plant cells directly sense water potential, we searched for D2O-dependent condensation by expressing proteins with the potential to undergo phase separation18 in cells and challenged the cells with an isosmotic buffer prepared with D2O. From this screen, we discovered a few proteins that formed condensates in tobacco cells in response to D2O treatment (Supplementary Table 1). Among them, At5g23680, which encodes a sterile alpha motif (SAM) domain-containing protein named SAM8, formed nuclear condensates in tobacco and yeast cells on D2O treatment (Fig. 1b,c). We next generated SAM8 complementation lines (SAM8pro::SAM8-mVenus) in sam8-1 knockout plants (Extended Data Fig. 3b–d). In root tip cells, SAM8-mVenus was diffuse in the nucleoplasm but formed nuclear condensates when plants were transferred into an isosmotic buffer containing more than 40% D2O for 5 min (Fig. 1d, Extended Data Fig. 1c,d and Supplementary Video 1). D2O-induced condensation of SAM8 was confirmed by immunostaining (Extended Data Fig. 1e). As a comparison, two known phase-separating proteins SEUSS and DCP511,12, did not form condensates on D2O treatment (Extended Data Fig. 1f). These observations indicate that the response of SAM8 to D2O is genuine and specific.
We next tested whether SAM8 condensation is responsive to water-potential change induced by hyperosmotic stress. SAM8 rapidly formed condensates in tobacco epidermal cells (Extended Data Fig. 1g), yeast cells (Extended Data Fig. 1h) and Arabidopsis root tip cells (Fig. 1e) when cells were treated with hyperosmotic stress. The threshold of water-potential change for SAM8 condensation was approximately 0.3–0.35 MPa (Extended Data Fig. 1i,j). The condensates are highly dynamic as they dissolved within 20 min on stress removal (Fig. 1e) and exhibited fast recovery of fluorescence after photobleaching (Extended Data Fig. 1k). Ethylene glycol (EG), which does not cause molecular crowding but reduces cellular water potential12, triggered SAM8 condensation (Fig. 1f), indicating that SAM8 condensation is not dependent on molecular crowding. Treatment with abscisic acid (ABA) and reactive oxygen species (H2O2), both of which are early signals in osmotic signalling, did not induce SAM8 condensation (Extended Data Fig. 1l). Moreover, chemical inhibition of calcium influx by LaCl3 or genetic ablation of Raf-like kinases (ok130-null mutant)19 had negligible impact on SAM8 condensation (Extended Data Fig. 1m,n). These results indicate that SAM8 condensation is independent of known osmotic signalling pathways.
The processes of seed ripening and germination experience marked changes in cellular water potential (Fig. 1g). SAM8 is highly expressed in seed and root tissues (Extended Data Fig. 3a), prompting us to examine SAM8 condensation during seed development and germination. In the course of seed development, when the embryos are still wet, SAM8 was diffused within the nucleus (Fig. 1h). Concomitant with dehydration during the late stages of seed development and in dry seeds, SAM8 formed condensates (Fig. 1h, i). On imbibition in water but not in NaCl solution, SAM8 condensates rapidly dissolved within 10–20 min (Fig. 1i). When water-imbibed seeds were transferred to NaCl solution, SAM8 condensates reappeared (Fig. 1i), suggesting that SAM8 condensation dynamically responds to water-potential changes in seeds.
Temperature also tunes cellular water potential5,20. It has been reported that temperature increases and decreases antagonize and synergize with hyperosmotic treatments, respectively, in terms of cellular water potential1. Indeed, we found that hyperosmotic treatment at 35 °C induced fewer SAM8 condensates than at 25 °C, whereas treatment at 15 °C markedly enhanced its condensation in yeast cells (Extended Data Fig. 1o,p). Statistical analysis showed a significant interaction between external osmotic strength and temperature for SAM8 condensation (Extended Data Fig. 1p), indicating that temperature and osmotic stress jointly regulate SAM8 condensation. Similarly, in Arabidopsis roots, although hyperosmotic treatment at 25 °C triggered SAM8 condensation within 5 min, the same osmotic strength at 37 °C failed to induce SAM8 condensation even within 20 min (Extended Data Fig. 4a).
Taken together, these data indicate that SAM8 undergoes water-potential-dependent condensation in cells.
The driving force of SAM8 condensation
SAM8 contains an SAM domain at its C terminus21 (Fig. 2a and Extended Data Fig. 2a). The remaining regions were largely unfolded (Extended Data Fig. 2a) and predicted as intrinsically disordered regions (IDRs; Fig. 2a). The SAM domain is a classic oligomerization domain that undergoes head-to-tail assembly and was reported to mediate phase separation22,23. Sequence-based predictions indicated that three positively charged amino acids (280 R, 281 R, 282 K) in the end helix surface are important for SAM8 oligomerization (Fig. 2a and Extended Data Fig. 2b). In vitro purified SAM domain mainly existed as oligomers, but substituting the three RRK residues with negatively charged glutamic acid residues (SAMRRKm) converted SAM to a monomer (Fig. 2b). To investigate the contribution of each region to SAM8 condensation, we generated truncation or mutation variants and tested their condensation in tobacco cells. Deletion of IDR1 resulted in partial cytoplasmic distribution of SAM8 but retained osmotic-dependent condensation, whereas deletion of IDR2 abolished SAM8 condensation (Fig. 2c). SAM8 with the SAM domain deleted (SAM8∆SAM) or mutated (SAM8RRKm) failed to form condensates (Fig. 2c). In line with this, co-expression of SAM8∆IDR2 and SAM8RRKm is sufficient to reconstitute osmotic-dependent condensation (Extended Data Fig. 2c). The necessity of IDR2 and SAM domain in SAM8 condensation was further confirmed in yeast cells (Extended Data Fig. 2d). In Arabidopsis stable transgenic plants, SAM8∆IDR2 and SAM8RRKm failed to form condensates on mannitol (Fig. 2d) or D2O (Extended Data Fig. 2e) treatment. These results indicate that both IDR-mediated multivalent interactions and SAM-mediated oligomerization are required for SAM8 condensation.
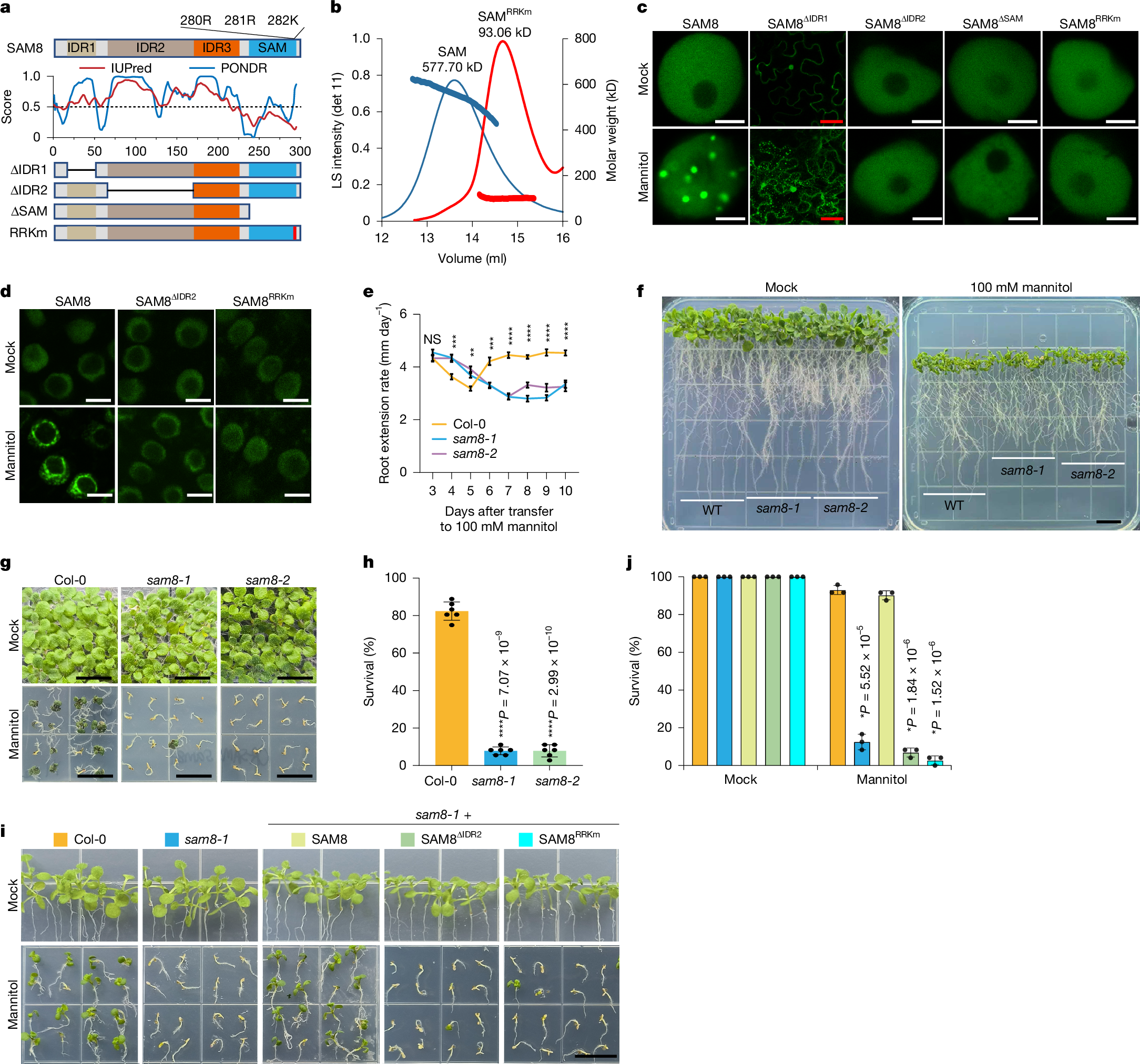
a, Top and bottom, domain structures of SAM8 and its variants. IDR, intrinsically disordered region. Middle, IDR prediction by the indicated algorithms. b, Size-exclusion chromatography coupled with multi-angle laser light scattering assay showing the size of the SAM domain and SAMRRKm. Both proteins were fused with an MBP tag to prevent misfolding. Left y-axis, the light scattering intensity from the 90° detector (detector 11). Right y-axis, molecular weight. c, Representative confocal images of tobacco epidermal cells expressing SAM8 and its variants. The cells were treated with or without 0.3 M mannitol. Scale bars, 5 µm (white); 50 µm (red). d, Representative confocal images of Arabidopsis root tip cells expressing SAM8 and variants treated with or without 0.3 M mannitol for 10 min. Scale bar, 5 µm. e, Quantification of the root extension rate of Arabidopsis seedlings that were transferred to medium supplemented with 100 mM mannitol. Error bars indicate mean ± s.e. (n = 30). Statistical significance details are provided as Source Data. f, Representative images of Arabidopsis seedlings that grew for 10 days after transferring to medium supplemented with or without 100 mM mannitol. Scale bar, 1 cm. g,i, Representative images of 12-day-old seedlings of the indicated genotypes grown on medium containing 300 mM mannitol. Scale bar, 1 cm. h,j, Quantification of the survival percentage shown in g and i, respectively. Error bars indicate mean ± s.d. (n = 6 replicates with each replicate containing 36 plants for h; n = 3 with each replicate containing approximately 24 plants for j). For e,h and j, asterisks indicate significant differences (two-tailed t-test). NS, not significant. For c and d, n = 3 independent experiments.
SAM8 condensation is functional
We next assessed the functional significance of SAM8 condensation. We obtained two mutant alleles: sam8-1 contains a T-DNA insertion that abolishes SAM8 expression (Extended Data Fig. 3b–d); sam8-2 is a CRISPR mutant with a 1-bp insertion that causes a premature stop codon (Extended Data Fig. 3b). Under normal growth conditions, sam8 mutants were phenotypically indistinguishable from wild-type Col-0 (Fig. 2f,g). When plants were exposed to hyperosmotic stress, both root apical meristem cell growth and root extension rate were inhibited in Col-0 during the first few days and partially recovered later (Fig. 2e,f and Extended Data Fig. 3e–h), whereas this inhibition and recovery were significantly compromised in sam8 mutants (Fig. 2e,f and Extended Data Fig. 3e–h). This led to the hypothesis that impaired growth inhibition under osmotic stress increases stress sensitivity and possibly cell death. SYTOX Green staining showed substantially higher cell death rates in the sam8 mutants than in Col-0 (Extended Data Fig. 3i,j), and the survival of sam8 seedlings under hyperosmotic stress was significantly reduced (Fig. 2g,h). The hypersensitivity of the sam8-1 mutant can be fully rescued by wild-type SAM8 but not by the condensation-defective variants (Fig. 2i,j). These data indicate that SAM8 condensation is essential for osmotic stress adaptation and tolerance.
In nature, hyperosmotic stress frequently coincides with higher ambient temperature or heat stress24. The dependence of SAM8 condensation on both osmotic potential and temperature (Extended Data Fig. 4a) prompted us to test whether SAM8 condensation mediates the combined effects. We found that hyperosmotic treatment at higher ambient temperature (26 °C) markedly increased death rates of wild-type plants compared with the same osmotic strength at 22 °C (Extended Data Fig. 4b–e). The sam8-1 mutant plants invariably displayed minimal root growth on osmotic treatment at both temperatures (Extended Data Fig. 4c–e). Consistent with this, when germinated and grown on hyperosmotic stress medium at two different temperatures, most of the Col-0 seedlings survived at 22 °C, but approximately 35% of them died at 26 °C (Extended Data Fig. 4f,g). The sam8 mutant plants showed no difference between 22 °C and 26 °C, and SAM8-complemented plants behaved like Col-0 (Extended Data Fig. 4f,g). By contrast, overexpression of SAM8 increased the propensity for condensation (Extended Data Fig. 4h) and enhanced plant survival at both 22 °C and 26 °C (Extended Data Fig.